P. Rotili , G. Gnocchi, C. Scotti, L. Zannone
Istituto Sperimentale per le Colture Foraggere - v.le Piacenza, 29 - 26900 Lodi (Italy)
1.Preliminary remarks
1.1 The breeding advances in forage yield in this century
Today, we have a good understanding of autotetraploid genetics and of its consequences in alfalfa breeding and variety synthesis. This was not true fifty years ago. In other words we have a good understanding of the mechanisms ruling heterosis and inbreeding effects. What has this new autotetraploid knowledge gained us? Certainly not much in the way of improving forage yield in itself (vigor). As alfalfa breeders have had only a limited success in increasing vigor they have turned much of their efforts to breeding for pest resistance, greater perennation and wider adptation. These pursuits have been highly successful.
It is true that the genetic improvement in forage yield for alfalfa in this century has been very low (about 10%). Why? There are the difficulties connected to the plant characteristics: autotetraploidy and allogamy. Other difficulties must be added: the reproductive apparatus with hermaphrodite flowers, the plant architecture (herbaceous plant) and most of all the meadow condition, that is density, numerical and biological. All that is not enough to explain the very moderate success obtained in breeding for yield. Other factors can play a role: for example, the research programme strategy and its effectiveness. In perennial species the groups studying the variety construction process are very rare. On the contrary, researches on the different phases of such a process are very numerous. Without a conceptual apparatus integrating the different phases in a unique programme, the great majority of these reasearches are very often repetitive and not significant. Today there are on one side excellent studies on molecular biology, genetics, physiology, etc.; on the other side the variety construction process is the same of fifty years ago. This situation of separation does not permit any progress and is very expensive.
Many people make varieties; among them, breeders who improve the varietal process by means of new ideas and discoveries are very rare. In the breeding work, genetics, physiology, agronomy, molecular biology, phytosociology, etc. play a part; however, breeding is not the sum but the integration of these subjects. Can breeding be defined as a science or a technology? It is both a science and a technology together. It is necessary to say that in the laboratories of science applied to agriculture the number of pathologists, biochemists, geneticists, cytologists and over all molecular biologists is increasing. On the contrary, the figure of the breeder has nearly disappeared. In a complex forage species as alfalfa it will be impossible to achieve important advances without breeding science. Important concepts as genetic load, biological density, adaptative reaction, vigor, linkat, level of complementarity and others will be able to show the whole of their operational value only through breeding.
1.2. Definition of vigor
Breeding for vigor includes all the most important traits and processes of the plant: forage yield, vegetative growth, resistance to various stresses or diseases, seed production and so on. The term vigor is therefore considered as a big container in which all our activity can be put.
If so, this term has no informative value: on the contrary, it can be confounding if its meaning is not clearly defined. In the glossary of genetics and cytogenetics this term does not exist. There is, of course, the term heterosis, proposed by Shull on 1940 during a Conference at Göttingen. Heterosis is the short form for heterozygosis; Shull used it to express the heterozygotic stimulation of which the hybrid vigor is a manifestation. Practically, the term heterosis is almost always utilized as a synonimous of "hybrid vigor". According to Shull, heterosis is the superiority of heterozygous genotypes with respect to one or more characters in comparison with the corresponding homozygotes. On the contrary, speaking of breeding for vigor, we use this term without any specification as a synonymous of productive value. We propose the following definition: vigor is the biomass productive capability of a genotype in the observed environmental conditions and in a defined time interval. This definition is valid for the cultivated plants. On the contrary, for wild populations, we consider the vigor as synonymous of reproductive value or fitness.
1.3. Estimate of vigor
The vigor is a property of the genotype as a whole, and it is more than the sum of the values of its single genes. Vigor is a concept and as a concept is not measurable. It is possible to estimate the vigor by measuring different traits. But the total dry matter yield, being the result of the activity of the whole genome, can be considered the best estimation of vigor.
There are many different criteria to estimate vigor by dry matter yield. Among them, the most precise is based on the following rule: the vigor of a genotype in a breeding program must be estimated in pedoclimatic and management conditions as close as possible to those in which the future variety will be utilized.
Therefore, first of all a clear definition of the type of variety to be constructed is needed: for instance, an alfalfa for grazing, for cutting, for dehydration, etc.
But it is not sufficient. It is necessary then to state how to measure the dry matter yield of the plants. It is generally assumed that in order to know all the productive potentialities of a plant we have to study it in non-limiting conditions, so that as spaced plants. In fact, the procedure of spaced plants is normally employed. But for the breeder it is not important to know the yielding possibilities of an alfalfa plant in non limiting conditions: for the breeder it is useful to know the yielding capacities of the alfalfa plant related to the alfalfa crop, that means as an element of the alfalfa crop system. In the alfalfa crop there is at least one limiting factor that can not be eliminated: the light. This factor constitutes perhaps the main difference between spaced plants and alfalfa crop.
At the surface of the alfalfa crop, the light is never a limiting factor; but when the height of the plants reaches 70-80 cm, at their base and in the lowest layers of the canopy (20-30 cm), the intensity of light decreases till 500-1000 lux, with effects on the senescence rate and persistence of the leaves in the first 40 cm, and also on the stem number at the regrowth. One of the most important task in the breeding for vigor is to invent the technical solutions allowing to simulate at the best the conditions of the plant in the different types of alfalfa crop. These techniques are much more effective than the traditional ones in the practical selection work and, in addition, allow a better estimation of some genetic parameters.
2. On the method
2.1. The elements of breeding schemes
Our research activity is aimed at the creation of new populations. Therefore, opinion on this activity is referred to a) the final product (the new variety) and b) the adopted process. For the first point, the aspects concerning forage production, quality and seed production; as for the second point, all those elements belonging to the subject called "breeding methodology" can be taken into account. The studies in this sector aim at linking the different phases of the new cultivar construction process. The breeder is not, as wrongly supposed, a variety maker but a scientist studying how to create a variety. Obviously, the verification of the validity of such a process must be the final product, i.e. the variety. Therefore, the breeder creates the variety prototype which will be a reference point for the new variety making activity. Breeding is the science of variety construction. This science is the result of the integration of different subjects such as genetics, agronomy, biology, phytosociology, etc. A breeding scheme is the way to represent the construction process of a biological product named cultivar. The variety construction process consists of three elements: the method, the procedure, and the techniques. The method is the series of successive steps and acts needed in the construction process of a new variety (analysis of parental populations - phenotypic selection - genotypic selection - synthesis); the procedure is the way to perform the method, i.e. the rules of the method carrying out. Finally, the techniques are the different ways of operating during the work.
2.2 Evolution of the traditional breeding schemes
Two typical breeding schemes, proposed in different periods, are now at disposal of the breeders for the construction of new cultivars. The comparison of these schemes allows us to follow the evolution of the ideas they are based on and the practice they use. Simplifying, the first scheme comprises the following successive methodological steps: a) phenotypical analysis of parental populations; b) phenotypical choice of parental plants; c) synthesis of new populations. This scheme was improved introducing the assessment of the genotypic value of the plant before the synthesis of the new population. We have therefore a second scheme characterized by the use of the progeny test which can be realised by progenies from polycross, top-cross, etc. The improvement concerns only the method. There is no difference between the first and the second scheme at the level of the procedure used. In fact, the spaced plant nursery remains the procedure normally adopted in the evaluation of individual plants and progenies. For the history of breeding, polycross was developped independently by H.N. Frandsen in 1940, by Tysdal in 1942 - who suggested the name "polycross" - and by Wellensiek in 1947, who used the term "mass test cross".
The value of the polycross depends on a real panmictic mating. From the theoretical point of view, the validity of polycross is unquestionable, both for the parental plant testing and for the construction of a synthetic variety. In practice, the suiility of polycross is very poor because the probabilities of having in the field a true panmixy are few. The chief factors producing an uncorrect panmixy are:
a) the entomophilous pollination. The effectiveness of different insects as alfalfa pollinators is very variable: there are good pollinators, insects causing the flower self tripping without pollination and insects making a selection based on flower colour and volatile compounds;
b) the difference in amount and quality of the parental pollens;
c) the reciprocal effects for pod fertility and seed setting ( 31, 35);
d) the different level of resistance to flower self tripping of the parents;
e) the different level of self fertility of the parents.
A polycross procedure performed by hand pollination without emasculation, with isolators allows to eliminate a part of these negative factors. But only the diallel cross ( by hand without emasculation) allows us to know the performance of the parents.
2.3. Reduction of heterozygosity
2.3.1.Selfing
Many researchers have studied the theorical basis of consanguinity and evolution of vigor during the selfing phase in autotetraploid species (2, 7, 10, 12, 16, 19). "Determining the effectiveness of inbreeding and subsequent selection at the inbred level to improve the genetic worth of parents is one of the more important objectives of alfalfa geneticists and breeders". So Busbice, Hill and Carnahan (6). We say that such an objective is surely the most important for a breeder. Why? Because it increases the effectiveness of selection . Our experimental data showed that selection in S0 and S1 was ineffective (24). We explained such results by the following hypothesis: the role of hybrid vigor masks the vigor due to the quality of genes and linkats. Selfing, by reducing heterozygosity (unmasking), allows the role of genes and linkats to emerge and makes possible the selection. For that reason, we consider 2-3 generations of selfing to be necessary.
In the course of years, we selfed more than 25.000 plants . The traits we studied on individual plant basis were: dry matter yield, height, stem number, earliness of regrowth and flowering, seed setting and pod fertility.
Our experiments were realized at the density of 4 - 150 - 500 plants / m2 (22, 23, 32, 36). The effects of selfing on morphological traits changed according to the density and the number of cuts when these last were made at the same time for all the material. The selfed material was slower in regrowth and flowering and therefore it cumulated through the cuts a delay in the restoration of the root reserves in respect of the S0; this delay brougth to a progressive loss in vigor and finally to an increase of mortality.
In the forties and fifties the inbreeding depression data have given rise to many discussion on the ploidy level of alfalfa. In general, in spaced plants, the trend of inbreeding effect on vigor did not follow the theoretical values expected in an autotetraploid, as set forth by Bartlett and Haldane (1), but it was much closer to the pattern expected in a diploid species.
From these and other observations, Wilsie (40) suggested that " a complex character such as vigor, conditioned by many genes and perhaps blocks of genes, may be inherited on a disomic basis".
Our opinion on this point is different. By varying the experimental conditions (either density or cutting regime, or arrangement of progenies in the field) we obtain different results of inbreeding depression (Table 1). The trend in vigor drop may in fact follow either the theoretical values expected in autotetraploids or those expected in diploid species, or other trends.
In short, the inbreeding effect on vigor (estimated as dry matter yield) cannot give an answer to the problem concerning the ploidy level of alfalfa. On the contrary, the recovery of vigor can give a sure answer on the ploidy level in any density condition, when we use partly inbred material. In fact the (Syn 2)S2 is always over the (Syn 1)S2. This surely demonstrates that vigor is inherited on a tetrasomic basis.
The principal advantages of selfing are:
a) it increases the effectiveness of selection;
b) it makes possible to homogenize the plant material for physiological traits. Large genetic variability for traits such as the earliness and quantity of regrowth, the growth rate and the earliness in flowering is a factor against persistence and forage yield stability. Such a variability in fact leads to different degrees of root reserve recovery at cutting time, with consequences on yield and persistence;
c) selfing assisted by selection is the most effective way to concentrate the genetic structures (genes and linkats) favorable to vigor. Therefore, it is possible to improve the breeding value of the parents (their General Combining Ability);
d) selfing is the most effective way to explore the genetic load;
e) it is possible to create plants "tolerant" to inbreeding.
2.3.1.1. Variability in inbreeding depression
There is a large variability among individuals for the inbreeding effect on vigor. Does this fact introduce the possibility of selecting for the "tolerance" to selfing? In addition, the traits weight, stem number and height show a different sensibility to selfing. The variability in inbreeding depression could be explained by many factors:
a) a variability in the level of heterozygosity of the parental material;
b) a variability of the effects of genes controlling either the root reserve dynamics, or the velocity of growth or the earliness in flowering. Inbreeding depression in a plant ranges from a maximum when all loci are tetragenic, to a minimum when all loci are monogenic. Question: do totally tetragenic or totally monogenic individuals exist? For many reasons we can say that it is possible but not probable. On the totality of individuals the rate of tetragenics and of monogenics is probably very lower than the rate of trigenics and digenic duplex and simplex. If so, inbreeding depression from S0 to S1 is linked to the nearly total disparition of tetragenics and to the reduction of trigenics. Passing to S2 tetragenics are reduced to 0.03 % and trigenic to 0.44 % (7). In short, variability in inbreeding depression is a consequence of variability in heterozygosity level. So the expression "tolerance" does not represent the reality. By contrast, the expression "sensibility" to selfing of the different traits is acceptable. It is possible to create by selfing the "tolerance" to selfing.
c) Another hypothesis: a variability in the organization of parental plants in genes and linkats (linkat number and dimension) implies a variability of response to selfing.
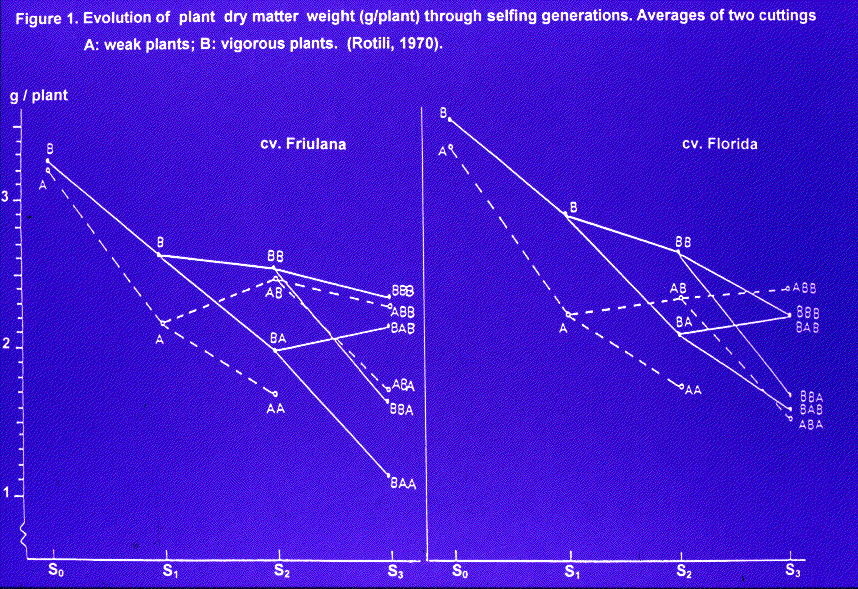
Going back to the problem of selection: is a selection effective when based on inbreeding depression? It is not effective because the comparison S1 vs S0 may be biased by the fact that S0 is the result of a cross S0 x S0 which could increase or not the rate of heterozygosity and therefore the vigour. So, it is possible that two S1 progenies show a same inbreeding depression on the corresponding S0, despite a dry matter weight very different. The selection has to be based on the vigor of the selfed progenies. Choice of the best progenies and of the best plants within the best progenies. Our results showed that selection of the best plants in the best progenies reduced inbreeding depression. (Fig. 1) At that time, we could not know if selection (in agronomic density condition) simply maintained the original heterozygosity or increased the frequency of favorable genes and linkats. Passing to the hybridization phase, experimental results showed that selfing associated to selection improved the genetic value of mother plants.This is the most important target of geneticist and breeder. Today, we can affirm that selfing associated to selection at agronomical density is a very powerful tool in order to improve the alfalfa genome and the stand structure.
2.3.1.2. Some remarks on selfing
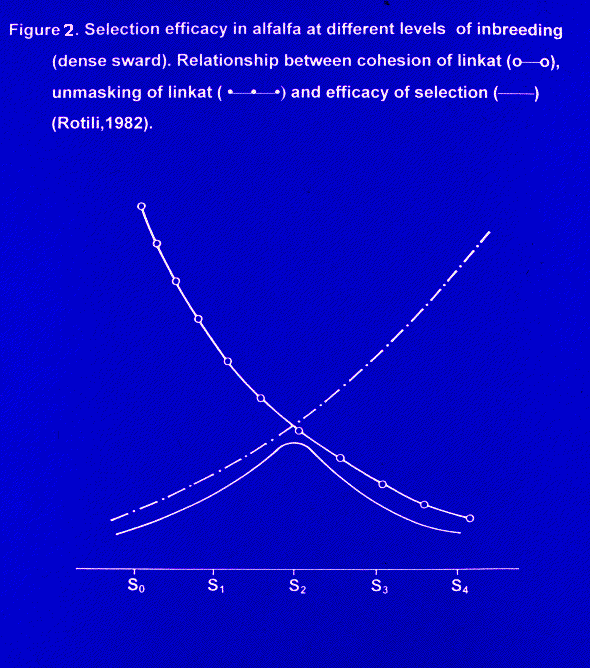
a) The linkat hypothesis is the only one allowing us to understand the rapid success of selfing in an autotetraploid plant. Let us admit that the alfalfa genome is made by genes and linkats. The genic content of linkats passes intact untill S2 - S3 generations. Does the linkat value equal the sum of genes or is it the result of a "system effect" bringing the linkat value over the sum of the single values of composing genes? Bingham et al. (3) give a different interpretation of linkat. For these Authors, the linkat is a linkage block of dominant alleles apt to create a complementary gene interaction process. Demarly (11) proposed the notion of "linkat", defined as a group almost stable of genes, constituting a coordinate whole having a great selective advantage. The linkat should have a local control system of the internal recombination rate that protects the linkat cohesion (named tension of linkat). This hypothesis, by postulating a hierarchic chromosomal organization, could explain the rapidity of concentration of factors favourable to vigor observed after a short phase of selfing. As the selfing level increases, the linkat unmasking increases too, while its tension decreases. The effectiveness of selection in an autotetraploid should be conditioned by these two facts (27).Selfing plays a positive role because, by reducing the heterozygous structure, it allows the selection to catch the linkats. But if selfing is pushed over a given threshold, the positive effect is cancelled because the linkats, losing in tension and therefore in stability, escape selection. The optimum seems to be at the second generation of selfing (Fig. 2).
These considerations bring to the following question: is the breaking of linkats useful for the variety construction and for productivity? We think more advantageous to preserve them and to undertake to research the selection techniques more suitable for their valorisation.
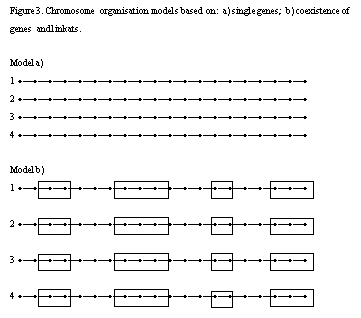
b) The scheme of Figure 3 shows two models of chromosome organization. We make the hypothesis that the a) scheme represents the chromosomal organization at the species origin; the b) scheme represents the organization of chromosomes after the action of natural and unintentional anthropic selection. This last model, that we can call "Demarly's model", is based on the coexistence of linkats and individual genes. The linkat length and internal cohesion can vary. The decreasing structural cohesion of linkats in S2 - S3 is due to the fact that selfing breaks off the genic interactions acting in trans. The same result can be obtained by the in vitro culture. Sibi et al. (38) made the hypothesis that genetic factors controlling the linkat cohesion act in trans for the allogamous species and in cis for the autogamous species. In conclusion, we can say that the genome value is represented by the quality of genes + quality of linkats + interactions within chromosomes + interactions between chromosomes (heterozygosity) .
c) In alfalfa the selfsterile plants are rare. Variability in autofertility is very high. Despite the lack of experimental results it is reasonable to presume that in selfing a competition within autopollen takes place. Such hypothesis is justified by the existence of a large variability among pollen grains for the average heterozygosity degree. As in the plants, we can speak of hybrid vigour of gametes. Such mechanism would maintain stable the level of heterozygosity in the individuals after the second and third selfing generations. We observed that statistically the very vigorous plants having a very high self fertility show a S1 and S2 progeny more performing compared with highly vigorous plants having a very low self fertility. It could be said that in the first case plant vigour is due to gene and linkat quality, while in the second case it is linked to the high heterozygosity level. In conclusion, we could say that self fertility has been created by natural selection starting from the plant autosterility.
2.4. Reconstruction of heterozygosity
2.4.1. Progeny test
We should consider the progeny test as the first step in the reconstruction of the heterozygosity when the material is partly inbred. It is to underline that in alfalfa it is impossible to obtain totally inbred individuals because of the reasons previously said. The recovery of the initial heterozygosity level can be realized at a different crossing generation succeeding the first one, according to the number of parents involved and to the degree of allelic diversity within the parental pool. This was experimentally demonstrated by several Authors (4, 15, 18, 24, 33).
Normally, the progeny test is made by the polycross. We said before that on theory the polycross progeny test is perfect, while in practice it can bring to great errors of evaluation. We also said that to minimize the effect of nonrandomness in polycross test we suggest to produce the F1 and F2 generations by hand. But if we want to know the genetic value of a parent the best solution is the diallel cross. Such a solution allows to know not only the combining abilities of parents but the reciprocal effects as well, very important in the evaluation of the fertility level of parents and therefore of their ability to produce seed (31, 35).
The results concerning vigor of diallel crosses S0 x S0 , S1 x S1 , S2 x S2 , S3 x S3, grown at a density of 330 plants/m2 (24, 31, 33, 35) suggest some answers to some crucial questions concerning alfalfa breeding: a) GCA was considerably greater than SCA at every level of selfing. Thus the greatest progress will result from working with general combining ability estimates; b) theoretically, vigor of single crosses between autotetraploid parents will decrease as their degree of inbreeding increases. However in our study where selection was practiced, the mean of some single-cross families increased with inbreeding. This increase was transmitted to the Syn 2 generation of synthetics. This indicates that selection of vigorous plants within the vigorous progenies during the selfing phase was successful. Intentional selection in competitive conditions reduces the inbreeding depression. On the basis of selfing only it was impossible to state whether selection simply maintained heterozygosity or if it concentrated favorable genes and linkats. On the basis of our results it is possible to say that intentional selection of vigorous plants increased the frequency of favorable genes and linkats.
From these results two questions arise. First, why was selection in an autotetraploid plant such as alfalfa so successful for a trait as complex as yield? Demarly coined the word "linkat" to represent the chromosome segment that remains intact for more than one generation. This hypothesis, simplifying the general structure of the genome, allows to understand the rapidity of concentration of favorable genes and linkats present in the mother plant. It is necessary only to assume that factors in favour of vigor are held together within a linkat. The use of interplant interference within the family during the selfing phase is an other element that may increase the success of selection. Therefore to obtain a positive result in alfalfa breeding for vigor, the use of these three factors, "selfing, selection and agronomic density" should be combined.
The second question concerns the different reaction of plants to selection: a) from our results (24), some plants reacted positively to selection and other did not; b) after the second generation of selfing further selection became ineffective. The following answer seems to be valid. The plants whose vigor was due not only to the high degree of heterozygosity but to favorable genes and linkats as well, reacted positively to selection until the S2 level. In the succeeding generations, further progress in accumulation of favorable genes and linkats appeared to stop. An explanation may be found by analyzing fertility. It would seem that when a given threshold of homogeneity is reached, it becomes very difficult to select plants having both a good concentration of favorable genes and linkats for vigor and a good self-fertility as well.
Concerning pod fertility, the results of the first part of our researches are grouped in Figure 4.
The self- and cross-fertility observed in this study were similar to those reported by other Authors (4, 8, 9, 14). The material used in the study was obtained through selection for vigor during four generations of selfing. At the same time, all the vigorous plants showing a very low self-fertility were discarded for operative reasons; in other words, an indirect selection for self-fertility should operate at some extent.
After such statements, the following considerations can be made on the data just presented:
a) The general ability to give seed was in the clones greater than the specific ability, at every level of selfing excepted the S4 level. Thus, the greatest progress in cross-fertility of the clones will result from working with general ability estimates. In addition, the ranks of the female parent values were in good agreement with those of the average values: such a result indicated that the female parents were prevailing in the determination of the average fertility of clones.
b) The average values of cross-fertility of the clones decreased with the increasing level of parental inbreeding, while these same clones produced single cross families giving a forage yield which increased with the degree of inbreeding (24). In other words, selfing and selection for vigor produced a positive effect for forage yield; at the same time, the effect for seed production was negative, because of the loss of heterozygosity produced by selfing.
c) Concerning the single crosses, at every level of inbreeding the maternal effects were more important than the general ability to give seed, the S0 level excepted. As we have seen for the clones, for the single crosses as well the female parents prevailed in the determination of the average fertility.
d) The cross fertility of the single crosses decreased from S0 x S0 to S4 x S4. On the contrary, the forage yield values increased with the level of selfing (35).
The results for pod fertility in the second group of diallel crosses (31), with plants grown at the density of 400 plants/m2, can be synthetized as it follows:
a) General Combining Ability (GCA) was much larger than Specific Combining Ability (SCA). At the same time, we can observe some negative cases of SCA;
b) General Reciprocal Effects (GRE) were much larger than Specific Reciprocal Effects (SRE). This means that in the selection of single crosses the direction of the crossing is very important.
These results suggest that the polycross method in the construction of synthetic varieties could give misleading results or results of little efficiency for the seed and forage production. This method, even without the selective effect of pollinators, does not permit us to know the reproductive possibilities of an individual plant of the polycross group. Besides, the actual situation is very far from panmictic conditions because it is very frequent to have a high variability among parental plants for quantity and quality of pollen which influences the pollen competition.Therefore even in the construction of synthetic varieties a better solution is to utilize the diallel cross method realized by hand without emasculation.
2.4.2 Variety construction
2.4.2.1 Synthetic varieties
The study of the evolution of synthetic varieties made with partly inbred parents gave the following results (24):
a) with parents originated from two different sources, Florida and Friulana, grown at the density of 330 plants/m2, generally the 2-clone Syn 2 yielded less than the 4- and 8-clone Syn 2;
b) the performance of the 2-clone Syn 2 showed a close association with the general combining ability of parental clones at all levels of inbreeding. A good correlation was also evident between the single-crosses and their 2-clone Syn2;
c) at every level of selfing, the best 4-clone Syn 2 was derived from the same four clones, having the highest GCA . The highest yield was obtained from these four clones at the S2 level.
On the data concerning the inbreeding depression the following observations can be made:
a) at the S0 level vigor decreased from the Syn1 to the Syn 2 regardless of the number of parents: no 2-clone Syn 2 yielded more than its respective Syn 1. Some 4-clone and 8-clone Syn 2 from partly inbred parents yielded more in the Syn 2 than in their Syn 1 generation;
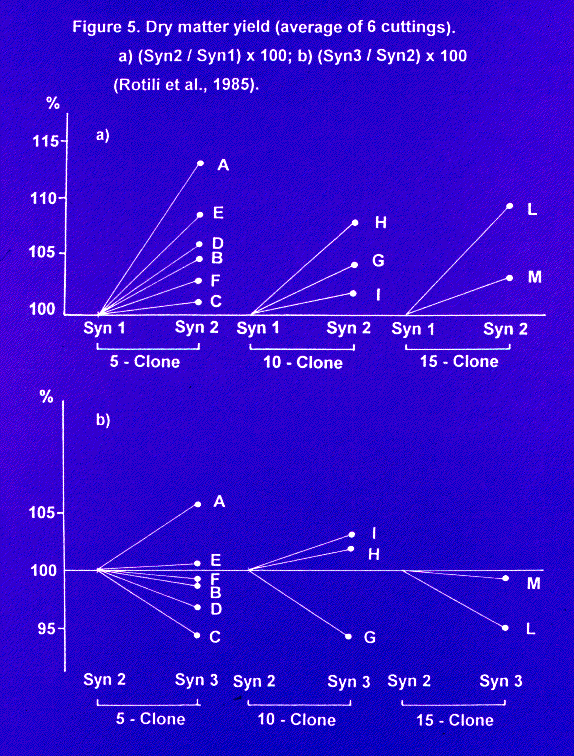
b) with parents originated from one source only, cultivar Lodi, grown at the density of 300 plants/m2 (29), a general increase in yielding capacity was observed from Syn 1 to Syn 2 generation (Fig. 5); by contrast, in Syn 3 several synthetics showed a decrease in yield when compared with their respective Syn 2. No effect of the number of constituents was observed. The trend from Syn 1 to Syn 2 is in agreement with the theory concerning the autotetraploids. The decrease in Syn 3 can be explained with the assumption that in Syn 2 the maximum level of heterozygosity is reached for some synthetics. The best synthetic variety in Syn 3 is based on 5 constituents.
The results in Syn 3 can be explained by different levels of allelic richness of the whole set of parental clones we utilized. When parental clones are partially inbred, an increased number of constituents may be not sufficient to prevent inbreeding depression in Syn 3 generation. Such cases of inbreeding depression in Syn 3 could perhaps be avoided by using clones derived from parental populations very different in genetic origin and growth area.
As for pod fertility, an increase was observed from the first to the second generation independently from the number of constituents of synthetics. In general, the average pod fertility increases with the increasing number of constituents, 5-clone synthetics showing a significantly lower fertility than 10-clone and 15-clone synthetics (Table 2).
2.4.2.2. Free-hybrids
Busbice (28) compares the coefficients of inbreeding in the "free hybrid" varieties with those of traditional synthetic varieties (Table 3). Interpreting the coefficients of inbreeding at face value and considering inbreeding to be deleterious, the synthetic varieties would always be superior to the free hybrids. However, this is no necessarily true. First, the computed coefficients of inbreeding assume random mating and equal survival of all offspring. We know that this does not occur. Inbreeding causes the death of many individuals. Different matings result in different levels of inbreeding and different rates of mortality. There is competition for survival among gametes, developing zygotes and plants. The result is that the computed coefficient of inbreeding are greater than the actual, and the larger the coefficient of inbreeding the greater is the difference between computed and actual values. Second, positive selection for desirable genes and linkats may occur or be practised by the breeder, during the phase of constructing varieties.
So, the value of the different methods of variety construction can not be determined from theoretical considerations alone. Rather, the methods must be evaluated experimentally. For the free hybrids to yield better than traditional synthetic varieties, the deleterious effects of inbreeding must be offset by positive selection for desirable genes and linkats during the selfing and synthesis phases of variety construction. Rotili and Guy (30) have chosen another way to compare synthetics at 4 and 8 components to the corresponding free-hybrid varieties (Table 4). The examined parameter is the number of alleles per locus and per plant. In free hybrids the calculation is based on the hypothesis of 100% crossing between the parental populations. Such hypothesis is not realistic. But even in the case of 50% selfing, the total number of selfed seeds would drop below 10% because of the negative effect of inbreeding on seed setting and pod fertility. This percentage is easily overcome by the "competition" effect at alfalfa stand level. Once these aspects cleared up, the advantage of free hybrid on the corresponding synthetic is of 0.33 and 0.18 alleles, for the hybrid with 4 and 8 parents respectively, at S2 level. Most recent data collected at Lodi during 3 years show that free hybrids with 4 S2 parents have a higher forage yield by 10% to 22% over the corresponding synthetics. But associated with the good forage yield there is a low seed production of parental synthetics (2 Syn 4) of free hybrid.
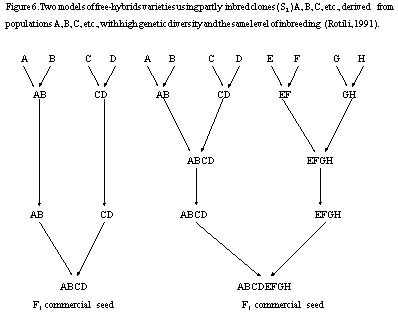
The program now in progress at Lodi, which will finish in 2002, is based on the comparison between synthetics at 4 and 8 constituents with the corresponding free-hybrids derived from 2 multiplied single crosses (double crosses) and from 2 multiplied double crosses ( octuple crosses) (Fig. 6). It is worthwhile to note that in previous experiments we observed that the best single crosses derived from the best S2 clones, and that the best double crosses derived from the best single crosses. Finally, we have to underline that in the case of octuple crosses, there are four different levels of intentional selection: during the selfing phase, at the level of single crosses, , at level of double crosses, during the multiplication of double crosses.
3. On the procedures
3.1. The importance of the procedures in the estimation of vigor and genetic parameters
The yielding capability of a cultivar becomes productive capability or vigor at the single plant level. The vigor is a property of the genotype as a whole, and it is more than the sum of the values of its single genes. The total dry matter yield, being the result of the activity of the whole genome, can be considered the best estimation of vigor. Studies on breeding of forage crops chiefly concern selection methods and variety models (5, 10, 11, 12, 13, 15, 16, 17, 20, 24, 25, 34, 39). There is another field not explored: it regards the plant with its biological environment constituted by other plants. Alfalfa cultivars are bred in programs based on spaced plants work and are grown as dense crop. We believe this is one of the most responsible factors for the obtained disappointing results. Our data show that either the choice of mother plants or the progeny test has to be made in dense sward . As previously said, infact, it is not important to know the vigor of an alfalfa plant in non-limiting conditions: on the contrary, it is useful to know the vigor of the alfalfa plant in relation to that of the alfalfa stand, i.e. as an element of the alfalfa crop system.
In a series of experiments different genetic materials (clones, polycross progenies, inbred families, diallel cross progenies) grown both as spaced plants and in dense sward were compared. The results concern different aspects of the problem: a) the prediction of forage yield in dense sward by observation of spaced plants; b) the importance of density, the plant arrangement and the material used (clones or progenies) for the estimate of the genetic parameters. Experimental conditions affect the choice of the plants. Our results showed a low correlation (r=0.4-0.6) between the spaced plants and the dense sward values for dry matter, plant height andnumber of stems. Besides, the correlation coefficients can change according to the different densities used. So, it is very important, when speaking about density, to define exactly the number of plants per square metre. Finally, we have to underline that cutting at 50% blooming does not impair persistence of spaced plants. On the other hand, in dense sward (300 plants/m2) the cutting effect is very selective, with about 20% mortality in the first year.
The estimate of genetic parameters is also affected. Our experiments showed that the value of heritability for dry matter yield and plant height varied with: a) density (spaced plants or dense sward); b) experimental procedures (pure stand, mixture in alternate rows or in the same row; c) the material used (clones or progenies); d) cutting management (cut at 50% flowering or at green bud stage; synchronic for the whole material or at a given biological stage for each clone or progeny). The same is true for the combining abilities. The use of spaced plants or dense sward modified the relative proportion of the different variances (GCA, SCA, Error) for dry matter yield (26).
Must the procedure of spaced plants then be completely discarded? In some conditions it can be efficient. In difficult pedoclimatic environments the forage production and chiefly the persistence are based on the stress resistance. Conversely, in easy environments production is due to the capacity of the plant to transform into biomass the naturally and artificially available resources. So, there are two different types of agriculture demanding two different types of biological machinery.The first has to resist to environmental stress; the other has to exploit the great resources of a rich habitat. The procedure of spaced plants can give positive results when hardiness has a predominant role; but it is ineffective in rich or non-stressing conditions, where the resistance factors play a secondary part in comparison with those directly involved in the expression of forage production. In fact, the factors of forage yield are not the same in spaced plants and in dense sward:
Spaced plants:
a) stem number; b) height; c) stem diameter.
Dense sward:
a) height; b) stem diameter; c) stem number.
In dense sward the cutting effect is very selective for the persistence of individuals.
3.2. Alfalfa crop and plant models
In practice, a single plant of alfalfa has no interest, because the exploitation concerns not a plant but the stand. We represent a stand as a system. In general a system can be defined as a set of elements plus the whole of the relationships among them that are globally called the structureof the system. In the alfalfa stand system the plants represent the elements; the structure is the morphological expression of the relationships among them. The aim of the breeder is to improve the alfalfa stand. A positive result is possible only if both the constitutive parts of the system are improved: the plants and the structure. Indeed, an optimal structure of the stand is necessary but not sufficient to secure a high forage yield. The same is true for the plants: a good genotype is indeed necessary but not sufficient.
Several alfalfa populations have been studied in association following an experimental scheme defined as "ecological diallel scheme". The relationship between partners found in alfalfa associations was always of "domination", improperly said competition (21). The cumulation of the negative effects of domination leads to a high overcompensation of the vigorous partners that results in the death of the weakest plants. In short, these experiments have produced the following results: a) annual and polyannual production is positively related to the degree of genetic homogeneity of cultivars; b) annual and polyannual mortality is negatively related to the degree of genetic homogeneity of cultivars; c) mortality is nearly absent in spaced plants but increases with density.
These results have important implications for the variety construction process: the most suitable model is a free-hybrid variety. The analysis of the interplant relationships makes clear that one of the most important objectives for the breeder is to improve the structure. This means to modify the relationships between individuals from a situation of domination (+,-) towards neutralism (0,0).
Some properties of the alfalfa stand producing the ideal structure ( monostratified structure) are:
A- Total synchrony of individual plants for:
a) earliness of regrowth; b) rhythm of internode elongation; c) earliness of the green bud stage.
B- Maximum homogeneity of individual plants for:
a) resistance to early cutting (at green bud stage); b) stem number and length of main stem after each cut; c) response to temperature and water supply through the different seasons.
The longevity of an alfalfa stand is not only the result of an improved structure but also of the degree of resistance of the plants to early cutting. A model of the ideal plant for a high density, frequently cut, irrigated alfalfa stand is presented:
a) plant dry matter:> X + 2 s;
b) leaf / stem ratio: 0.85-1.00;
c) average length of the three main stems at the first reproductive node:> X + 2 s;
d) stem number: about average;
e) stem thickness (diameter at the base of the main stem): 2.5-3.5 mm;
f) very high internode number of the main stem;
g) early and vigorous regrowth;
h) resistance to early cutting;
i) very positive response to summer temperatures and water supply;
l) early growth in spring;
m) fall semi-dormancy;
n) very high vigor combined with late appearance of the first reproductive node.
This two models have been set up through many experiments during the last 20 years.
4. On the techniques
A good technique must have at least two characteristics: first, to be suitable for the achievement of the fixed procedure; second, to allow time and money saving .
In our opinion, the more important technical aspects concern: a) the plant growth conditions; b) the by hand pollination; c) the test of heterozygosity level by molecular markers. As for the plant growing at a density comparable with that normally used in field conditions, we experienced a lot of solutions. The one we currently use is a PVC plot - tube 80 cm high and 5 cm diameter provided with an enlarged superior opening for watering. A second tube 3 cm diameter provided with a series of holes is put inside the outer tube in order to improve drainage. The single plant growing inside the inner tube is at a density equivalent to about 500 plants/m2; as for the aerial part the density effect is simulated by means of a shadowing net surrounding each tube. This technical solution makes possible a precise and quick evaluation of the plants and allows to move them handily wherever we want. This possibility is of basic importance when selfing and crossing are carried out.
We previously said that only the diallel scheme allows a correct evaluation of parental plants both as female and as male parents. The by hand pollination without emasculation requires a very high quantity of allopollen on the spatule. We verified that this technique assures a successful cross-pollination.
As for the molecular techniques, we propose the use of RFLP or other codominant molecular markers, for the estimation of heterozygosity and genetic diversity of the most vigorous plants in order to select the most performing individuals with a low heterozygosity level and with a high degree of diversity. In our variety construction process the first estimation concerns the selfing phase, the second one concerns the variety construction phase. Our results show that the number of bands revealed by RFLP technique decreases as inbreeding increases. This result is coherent with inbreeding depression measured on plant dry matter yield (37). It is worthwhile to notice that within different S2 families the very small plants show on average the same number of bands than the very vigorous plants. This should indicate that in S2 the difference between the two groups is due not to heterozygosity level but to the quality of genes and linkats. It is to note that both vigorous and weak S2 plants come from S0 high yielding parental plants.
5. For the future
A new alfalfa variety must have higher vigor and quality of forage and persistence to be succesfull on the market. But it is not enough; an outstanding seed-yield potential is needed. In other words the new variety must be a well-balanced product from the biological point of view. For this reason there is a very little probability that molecular biology and other biotechnological techniques may directly produce a population adapted to agricultural practices. From molecular biology we could get resistance sources to introduce in the variety constitution process. On the contrary the molecular markers seems to be the most promising sector; selection assisted by molecular markers (RFLP or other codominant marker type) during the process of variety construction may concern different fields:
b) the prediction of values of a cross through information on parents;
c) the management of variability at the genetic resources level sensu lato: grouping of clones or single hybrids based on combining ability.
Once that said, future researches should resume the studies on breeding methodology. Some crucial points seem to be:
a) to set up a plant model for vigor, quality (optimal leaves/stems ratio), for the resistence to a high number of cuts per year and for seed production;
b) to resume studies on selfing for breeding;
c) to set up a variety model allowing the production of F1 commercial seeds;
d) to find a molecular marker technique (RFLP or other codominant marker type) allowing to rapidly monitorize heterozygosity level and genetic diversity of a population;
e) to look for alfalfa transformation for the characteristics of the reproductive apparatus in order to have larger seed size, as in some annual species.
6. Operating plan of the Lodi Institute breeding methodology for the construction process of alfalfa commercial varieties
A - Choice of parental populations
Aa - Populations not related, with a good level of productivity and of desease and insect resistance; fall dormancy rating about 6.
B - Parental population study
Ba - Population growing at the density of about 400 pl/m2; cutting at 50% of flowering: 4-5 cuts during the sowing year, 2 cuts during the second year before selfing.
Bb - Traits to be evaluated per individual plant: dry matter weight; height at the first inflorescence of the main stem; number of stems (over 25 cm length); phenological stage, expressed by the number of inflorescences on the main stem.
Cb - Phenological stage at cutting: blue bud.
Cc - Stem morphology: short internode length; high internode number.
Db - Study of S1 plants (70 - 100 per family): as at paragraph B.
Dc - Choice of S1 plants within each family: as at paragraph C.
Dd - Selfing of S1 plants : as in Da.
De - Study of S2 plants (70-100 plants per family): as in paragraph B.
Df - RFLP or other codominant marker test of the most performing S2 plants.
Dg -Choice of S2 parental plants: as in paragraph C and taking into account the low level of heterozygosity estimated by RFLP or by other codominant markers.
Eb - Study of progenies: as in paragraph B.
Ec - Choice of the best parental single crosses within the series with
the highest GCA.
Fa.2 - RFLP or other codominant marker test of the 5 plants.
Fa.3 - Growing in the field of the maximum number of 2 Syn 2 plants of each parental single cross. Plant arrangement: rows cm 50 apart; 15 cm among the plants on the row. Homogeneization for the physiological traits in Fa.1.
Fa.4 - Construction in the field of double semi-hybrids with 4 constituents not related, [(ABxCD)] S2 taking into account the RFLP or other codominant marker test for genetic diversity. Arrangement in binary rows 50 cm apart, 15 cm within the two rows.
Fb.2 - RFLP or other codominant marker test of the 3 plants before the construction of the double crosses.
Fb.3 - Growing in the field of the maximum number of plants for each double cross following the indications in Fa.3.
Fb.4 - Construction in the field of the octuple semi-hybrids [(ABCD) x (EFGH)] with 8 S2 constituents not related taking into account the RFLP or other codominant marker test for genetic diversity. Arrangement as in Fa.4.
References
2 . Berninger, E. 1967. Contribution à létude des modalités dexpression de lhétérosis chez les autotétraploïdes. Ann. Amélior. Plantes 17:92-104.
3 . Bingham, E.T., R. W. Groose, D. R. Woodfield, and K.K. Kidwell. 1994. Complementary gene interactions in alfalfa are greater in autotetraploids than diploids. Crop Sci. 34: 823-829.
4 . Busbice, T. H. 1968. Effects of inbreeding on fertility in Medicago sativa L. Crop Sci. 8:231-234.
5 . Busbice, T. H. 1970. Predicting yield of synthetic varieties. Crop Sci. 10:265-269.
6 . Busbice, T. H., R. R. Jr. Hill, and H.L. Carnahan. 1972. Genetics and Breeding Procedures. p. 283-318. In Alfalfa Science and Technology ASA, CSSA, SSSA, Madison, WI.
7 . Busbice, T. H., and C. P. Wilsie. 1966. Inbreeding depression and heterosis in autotetraploids with application to Medicago sativa L. Euphytica 15:52-67.
8 . Dattée, Y. 1972. Analyse quantitative de lauto et de linterfertilité chez quelques familles de Luzerne. Ann. Amélior. Plantes 22 (I):5-21.
9 . Dattée, Y. 1974. Effet de lapparentement sur la production de graines en croisement chez la Luzerne (Medicago sativa L.). Ann. Amélior. Plantes 24 (I):25-35.
10. Demarly, Y. 1963. Génétique des tétraploïdes et amélioration des plantes. Ann. Amélior. Plantes 13:307-408.
11. Demarly, Y. 1968. La sélection des variétés synthétiques. p. 267-295. In Proc. V Eucarpia Congress, Milan, Italy. 30 Sept.-2 Oct. 1968.
12. Demarly, Y. 1971. Genetic comparison in di and tetraploids. p. 9-37. In Proc. of Eucarpia Fodder Crops Sect. Meet., Lusignan , France. 15-17 Sept. 1970.
13. Demarly, Y. 1977. Génétique et Amélioration des plantes. Ed. Masson, Paris, 1977, 287 p.
14. Dessureaux, L., and A. Gallais. 1971. Evolution of fertility in advanced generations of an Alfalfa single cross hybrid. Can. J. Genet. Cytol. 13:834-841.
15. Gallais, A. 1968. Evolution de la vigueur des variétés synthétiques tétraploïdes au cours des générations de multiplication. Ann. Amélior. Plantes 18:5-15.
16. Gallais, A. 1977. Contribution à létude théorique et expérimentale de lhétérosis chez une plante allogame autotétraploïde. Thèse Doctorat dEtat. Université de Paris, 341 p.
17. Guy, P. 1966. Etude de lévolution de quelques structures autotétraploïdes au cours des générations de multiplication. Genetica Agraria 20:56-65.
18. Guy, P. 1968. Nombre de plantes nécéssaires à la multiplication dun autotétraploïde. Ann. Amélior. Plantes 18:367-379.
19. Haldane, J.B. 1930. Theoretical genetics of autopolyploids. J. Genet. 22:349-372.
20. Hill, R. R. Jr. 1975. Parental inbreeding and performance of alfalfa single crosses. Crop Sci. 15: 373-375.
21. Jacquard, P. 1972. Glossaire de termes et définitions. Sect. Plantes Fourragères Eucarpia. Cepe, Montpellier, France, 36 p.
22. Rotili, P. 1970. Lautofecondazione nel miglioramento genetico dellerba medica. p. 5-69. In Quad. Sper. Ist. Colt. Forag.,1, Lodi, Italy.
23. Rotili, P. 1971. Selfing effects on S2 and S3 generations in Medicago sativa L. p. 169-188. In Proc. Eucarpia Fodder Crops Sect. Meet., Lusignan, France. 15-17 Sept.1970.
24. Rotili, P. 1976. Performance of diallel crosses and second generation synthetics of alfalfa derived from partly inbred parents. I. Forage Yield. Crop Sci. 16:247-251.
25. Rotili, P. 1977. Performance of diallel crosses and second generation synthetics of alfalfa derived from partly inbred parents. II. Earliness and mortality. Crop Sci. 17:245-248
26. Rotili, P. 1979. Contribution à la mise au point dune méthode de sélection de la Luzerne prenant en compte les effets dinterférence entre les individus. I. - Etude expérimentale de la structure de la luzernière. Ann. Amélior. Plantes 29:353-381.
27. Rotili, P. 1982. Considerations on some aspects concerning the breeding of meadow fodder species. p. 47-70. In Ann. Ist. Colt. Forag., 6, Lodi, Italy.
28. Rotili, P., T.H. Busbice, and Y. Demarly. 1996. Breeding and variety constitution in alfalfa: present and future. p.163-180. In Grassland and Land Use Systems. Proc. 16th EGF Meet., Grado, Italy. 15-19 Sept. 1996.
29. Rotili, P., G. Gnocchi, C. Scotti, and L. Zannone. 1985. Performance of second and third generation synthetics of alfalfa derived from partly inbred parents: forage yield and fertility. p. 135-141. In Proc. Eucarpia Fodder Crops Sect. Meet., Svalöv, Sweden. 16-19 Sept. 1985.
30. Rotili, P. and P. Guy. 1991. Breeding schemes in autotetraploid forages: recent achievements and new approaches. p. 14-22. In Proc. Eucarpia Fodder Crops Sect. Meet., Alghero, Italy.14-19 Oct. 1991.
31. Rotili, P., C. Scotti, D. Kertikova, and G. Gnocchi. 1998. Performance of diallel crosses of alfalfa with different levels of genetic diversity and derived from partly inbred parents. I. Seed setting and pod fertility. p. 48. In Rep. 36th NAAIC, Bozeman, MT, USA. 2-6 Aug. 1998.
32. Rotili, P., and L. Zannone. 1970. Caduta di vigore e competizione nellerba medica. p. 53-64. In Quad. Sper. Ist. Colt. Forag.,2, Lodi, Italy.
33. Rotili, P., and L. Zannone. 1974. General and specific combining ability in lucerne at different levels of inbreeding and performance of second generation synthetics measured in competitive conditions. Euphytica 23:569-577.
34. Rotili, P., and L. Zannone. 1976. Performance of diallel crosses among single crosses of alfalfa and comparison of double crosses and synthetics derived from the same parental clones. p. 55-66. In Proc. Eucarpia Fodder Crops Sect. Meet., Roskilde, Denmark. 7-9 Sept. 1976.
35. Rotili, P., and L. Zannone. 1977. Quantitative analysis of fertility in Lucerne at different levels of selfing. Ann. Amélior. Plantes 27 (3):341-354.
36. Rotili, P., L. Zannone, and G. Gnocchi. 1979. Effects of inbreeding on vigor of alfalfa measured in competitive and frequent cutting conditions. p. 111-119. In Proc. Fodder Crops Sect. Meet., Perugia, Italy.4-6 Sept. 1979.
37. Scotti, C., C. Suardi, F. Pupilli, F. Damiani, S. Arcioni, and P. Rotili. 1994. Inbreeding depression in Medicago Sativa: Estimation of the different levels of heterozygosity by RFLP. p. 182-184. In Rep. 34th NAAIC , Guelph, Ontario, Canada. 10-14 July 1994.
38. Sibi, M., M. Biglary, and Y. Demarly. 1984. Increase in the rate of recombinants in tomato (Lycopersicon esculentum L.) after in vitro regeneration. Theor. Appl. Genet. 68:317-321.
39. Theurer, J.C., and L. J. Elling. 1964. Comparative performance of diallel crosses and related second generation synthetics of alfalfa, Medicago sativa L. III. Forage Yield. Crop Sci. 4:25-28.
40. Wilsie, C. P. 1958. Effect of inbreeding on fertility and vigor of alfalfa. Agron. J. 50:182-185.
|
|
|||
|
|
|
|
|
| Spaced plants | |||
| (Tysdal et al.; Wilsie; Guy; Panella and Lorenzetti; Rotili and Zannone) |
|
|
|
| Dense sward (33 plants per linear metre; among lines distance 20 cm) |
|
|
|
| Dense sward and frequent cutting (66 plants per linear metre; among lines distance 20 cm) |
|
|
|
Table 2. Average pod fertility in Syn 1, Syn 2
and Syn 3 (Rotili et al., 1985).
| Number of parental clones |
|
|
|
|
| Generation of synthetics: | Syn 1 |
|
|
|
| Syn 2 |
|
|
|
|
| Syn 3 |
|
|
|
Table 3. Comparison of the coefficients of inbreeding
in the free hybrid varieties and in the traditional synthetic varieties
(Busbice, 1996).
| Variety models |
|
|
|
| Free hybrid | (S2 parents) |
|
|
| Synthetic | (S2 parents) |
|
|
| Synthetic | (S0 parents) |
|
|
Table 4. Comparison of 4 - and 8 - clone synthetic
varieties with free hybrids derived from 2 and 4 parental synthetics. Mean
number of alleles per locus and per plant (Rotili & Guy, 1991).
| Number of parental clones |
|
|
|
|
| Level of inbreeding | ||||
| S0 | Syn3 |
|
|
|
| free hybrid |
|
|
||
| S2 | Syn3 |
|
|
|
| free hybrid |
|
|
||
| S4 | Syn3 |
|
|
|
| free hybrid |
|
|
||
Table 1. Dry matter in percentage of non inbred.
Average of several trials (Rotili et al., 1979).
Table 2. Average pod fertility in Syn 1, Syn 2
and Syn 3 (Rotili et al., 1985).
Table 3. Comparison of the coefficients of inbreeding
in the free hybrid varieties and in the traditional synthetic varieties
(Busbice, 1996).
Table 4. Comparison of 4 - and 8 - clone synthetic
varieties with free hybrids derived from 2 and 4 parental synthetics. Mean
number of alleles per locus and per plant (Rotili & Guy, 1991).
Figure 1. Evolution of plant dry matter weight (g/plant) throuh selfing generations. Averages of two cuttings . (Rotili, 1970).
A: weak plants ; B: vigorous plants.
Figure 2. Selection efficacy in alfalfa at different
levels of inbreeding (dense sward). Relationship between cohesion of linkat
(o?o), unmasking of linkat (·?·?·) and
efficacy of selection (??) (Rotili, 1982).
Figure 3. Chromosome organisation models based on:
a) single genes; b) coexistence of genes and linkats.
Figure 4. Performance for pod fertility at different
levels of selfing (Rotili and Zannone, 1977).
Figure 5. Dry matter yield (average of 6 cuttings).
a) (Syn2 / Syn1) x 100; b) (Syn3 / Syn2) x 100 (Rotili et al., 1985).
Figure 6. Two models of free-hybrids varieties
using partly inbred clones ( S2 ) A, B, C, etc., derived from
populations A, B, C, etc., with high genetic diversity and the same level
of inbreeding (Rotili, 1991).