Gene transfer for alfalfa improvement
S. Arcioni, M. Bellucci, F. Damiani, F. Pupilli
Istituto di Ricerche sul Miglioramento Genetico delle Piante Foraggere del C.N.R.. Via della Madonna alta, n. 130, 06128 Perugia, Italy
Gene transfer is the possibility of incorporating wild and alien genes or manipulated sequences into a host genome and letting them express and pass down to the offspring. Alfalfa, as nearly all the species that have been cultivated for centuries could benefit enormously by this technology because natural variation has by now been largely exploited and the possibility of introducing superior traits through sexual crosses and selection is very limited.
While traditional plant breeding has given an invaluable contribution to the production of cultivated varieties in fodder crop species, productivity and plant performance are now making no headway for a number of concurrent reasons: the mechanism of reproduction and the interaction between additive and dominant components of variation, which are difficult to split apart because of the autotetraploid status of the species, the arising of intraspecific competition and the lack or silencing of genes for the expression of some useful compounds.
The introduction of new genes into the alfalfa genome may widen the field of application of traditional breeding, providing new traits and, through a modification of the method of reproduction, new strategies of variety breeding. Short, simple traits, such as pathogen resistance and production of secondary metabolites, but also more complex traits, such as plant shape and vegetative cycle, reproductive system and adaptation, could be improved by introducing alien genes into the M.sativa genome.
Two strategies have been followed in our lab for this purpose: somatic hybridisation and Agrobacterium transformation.
The work involved in somatic hybridisation can be summarised in three phases: 1) screening for suitable sources of M.sativa protoplasts; 2) setting up experimental conditions for protoplast fusion, heterokaryon selection, and regeneration of hybrids; 3) characterisation of somatic hybrid plants.
In order to take full advantage of the potential of somatic hybridisation as a tool for gene transfer in M.sativa, the genome of this species was combined with those of other Medicago species by protoplast electrofusion. Two of the hybrids were formed with closely related diploid species from the M.sativa complex, namely M.coerulea and M.falcata, both of which are capable of sexual hybridisation and gene exchange with M.sativa in nature when they are at the same ploidy level. The third somatic hybrid involved the distantly-related tetraploid shrub M.arborea which is incapable of sexual hybridisation with M.sativa and is the sole species of a separate section of the Medicago genus (McCoy and Bingham, 1988). We chose these three parental combinations to investigate separately the genetic consequences of somatic hybridisation between parents either at different ploidy levels or genetically distant. For practical applications, hexaploid somatic hybrids resulting from alfalfa and M.coerulea or M.falcata could represent useful biological material for the constitution of highly heterozygous varieties, able to overcome the inbreeding depression affecting alfalfa cultivars in stands characterised by poor pollinating conditions; M.arborea could be a source of genes for stress adaptation and winter forage production.
Early experiments of protoplast fusion involved, as parents, M.sativa cell suspension protoplasts and mesophyll protoplasts of other species, and although both parental genotypes were embryogenic, hybrid calli failed to regenerate.
Different factors preventing regeneration have been proposed: excess chromosomes (Arcioni et al., 1987), parental incompatibility, etc.. Nevertheless, through electrofusion experiments between protoplasts of the same embryogenic genotype and isolated from different tissues, we demonstrated that an epigenetic status arising in the cell suspension-derived protoplasts, correlated with a typical electrophoretic pattern of esterase and peroxidase, negatively affected regeneration of hybrid calli. As a matter of fact, the hybrid calli resembled for regeneration capability to protoplasts showing the highest plating efficiency. This series of experiments allowed setting up a strategy for the selection of parental protoplasts based on the idea that the protoplast source with the highest plating efficiency determines all the traits of hybrid calli, therefore the most in vitro active protoplasts must also be able to regenerate plants (Pupilli et al., 1991).
Major efforts were directed towards developing a method for the identification and selection of heterokaryons, which could be suitable for each parent combination involving M.sativa as recipient parent. Hybrid cells were selected on the basis of their colour, starting from parental protoplasts of different sources. Hybrids between green mesophyll protoplasts and colourless cell suspension or root protoplasts were identified on the basis of their
pigmentation (Fig. 1, A-D), half green and half clear (Damiani et al., 1988). Pupilli et al. (1991) improved this technique by treating colourless protoplasts with a fluorescent dye, extending (Fig.1, E-G) the period of hybrid identification by another 3-4 days, thereby reducing the risks of misinterpretations. Hybrid recovery was performed manually 2-3 weeks later after embedding the fusion products in a semisolid medium and recording the position of heterokaryons on the basis of their coordinates (Arcioni et al., 1987). This method allowed supplementing the culture medium with highly dividing parental protoplasts (Pupilli et al., 1992) that positively affect mitotic divisions of heterokaryons. Similarly, the surrounding of newly picked hybrid calli with actively dividing nurse calli markedly enhanced their growth.A crucial point in somatic hybridisation is the regeneration of hybrid plants. Combining the regenerating capacity of one parent with the high in vitro performance of the other parent has proved to be possible in some species, but not in Medicago. Therefore, as previously discussed, the genotype and tissue-source of parental protoplasts must be properly selected . However, even when parental protoplasts are properly selected, regeneration from hybrid calli remains difficult and slow (Arcioni et al., 1994). In fact, although M.sativa + M.coerulea somatic hybrids (S+C) followed the regeneration pathway of the sativa parental line, up to one-year's culture in the regeneration medium was needed to recover embryos from M.sativa + M.arborea (S+A) and M.sativa + M.falcata (S+F) hybrid calli (Arcioni et al., 1994). Quite possibly during such a long time parental genomes may have recombined and genomic rearrangements may have affected regeneration ability; as a matter of fact, hybrid calli have low regeneration efficiency than parents.
Studies on genome architecture, chromosome stability and field performance have been undertaken to ascertain the value of these plants. Here are reported the comparative analyses of the three somatic hybrids S+C, S+A and S+F carried out at cytological, molecular and morphological levels.
a) Cytological analysis
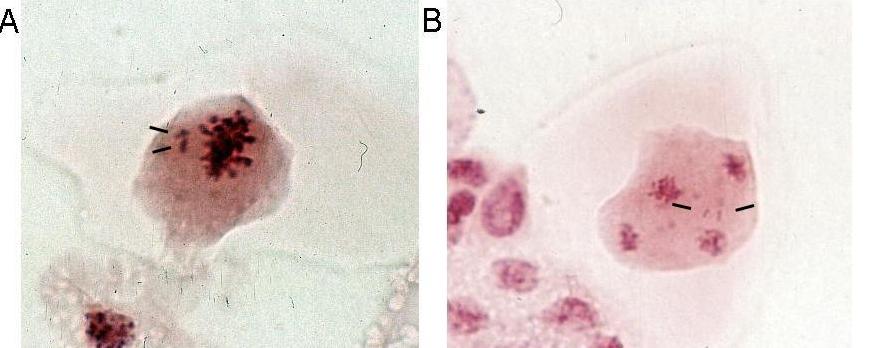
Chromosome counts showed that all parental chromosome sets were present in S+C. These hybrids had 48 (32 + 16) chromosomes and microsporogenesis analysis revealed minor meiotic abnormalities, such as essentially formation of univalents at metaphase I (
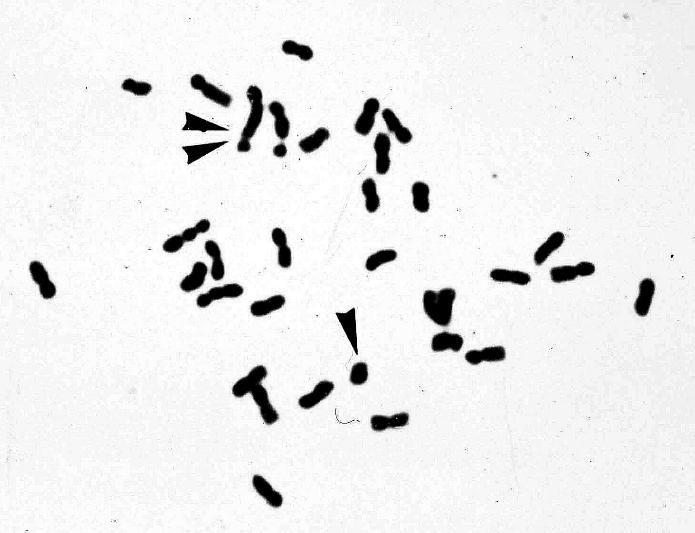
Fig. 2A) and lagging chromosomes at anaphase II (Fig.2B). However, meiosis appeared to be more stable than has been previously reported for spontaneous or cross-derived hexaploid alfalfa plants (McCoy and Bingham, 1991). A moderate loss of chromosomes was observed in S+A: these hybrids had 56 chromosomes, eight less than the sum of the two genomes (32+32). However, the uniform morphology of the parental chromosomes makes it impossible to determine the origin of the missing chromosomes. The S+F were hyperaneuploid (2n=33) and contained two extra-long chromosomes, indicating that a translocation had taken place (Fig.3).b) Molecular analysis
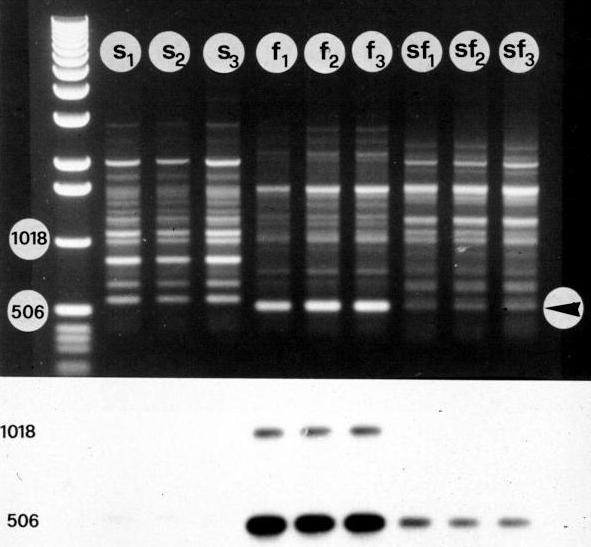
A molecular characterisation of the nucleus of S+C (Pupilli et al., 1995) was carried out on 15 plants derived from the same fusion event, by means of 27 genomic probes, 8 of which were located in the largest linkage groups of an alfalfa RFLP map (Brummer et al., 1993). No variation of the RFLP pattern was detected within the population of hybrids while some differences occurred between the patterns of the hybrids and those of the parents. Actually, about 30% of the M.coerulea specific bands were missing in the hybrids. These findings indicate that tissue culture did not play any role in the observed alterations, otherwise the long duration of hybrid calli culture would have introduced some variability into the hybrid population (
Fig. 4).In the S+A hybrids, approximately one third of the M.sativa-specific bands and half of those of M.arborea were absent from the pattern of the somatic hybrids, and 7 bands specific to the somatic hybrid plants were scored indicating large-scale genome rearrangements. Chromosome loss alone could not account for all the alterations observed: the appearance of new bands in the hybrid pattern indicated that genome rearrangements were also due to intensive mutational events, such as gene conversion, unequal crossing-over, gene duplication, etc., which could be responsible for any gain, or loss, of restriction sites and, consequently, for the lack of parental bands and the appearance of new ones. In S+F, the RAPD analysis was carried out mainly to determine if and how M.falcata contributed to the hybrid genome. Out of 22 random primers screened, nine produced at least one M.falcata-specific band present in the hybrids (
Fig. 5). The S+F RAPD patterns generally reflected the combination of the two parents while some parental RAPDs were lost. From Southern analysis two M.falcata-specific sequences were detected in the hybrid genome.More comprehensive data obtained combining cytological (Fluorescent in situ Hybridisation, FISH) and molecular (RFLP) tools were produced analysing the NOR (nucleolar organiser region). We selected this region because it contains thousands of tandemly repeated rDNA genes organised into one or a few loci where the highly-conserved coding sequences of adjacent repeat units are separated by highly variable regions (intergenic spacers, IGS), (for reviews see Long and David, 1980), which can change rapidly within somatic lineages (Rogers and Bendich, 1987) under particular stresses.
S+C retained all six parental NORs and all parental RFLPs representing a complete integration of rDNA (
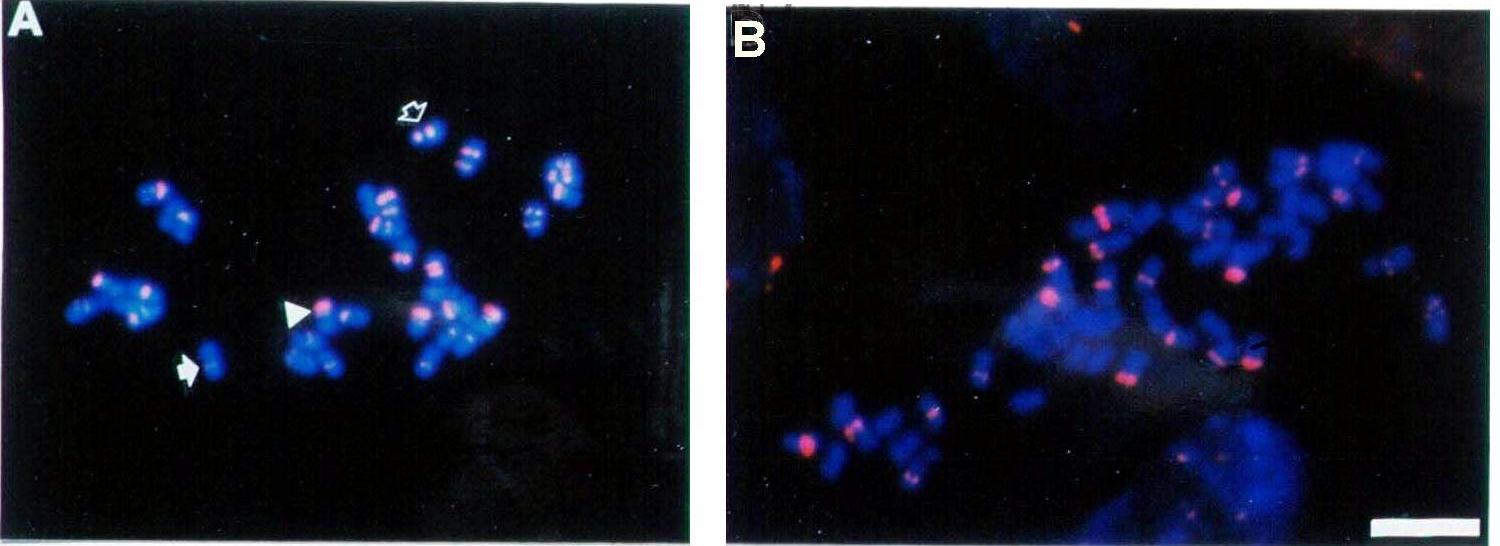
Fig. 6A). S+A retained five of six parental NORs while it lost half of the M.arborea-specific RFLPs indicating that simple chromosome loss of one M.arborea NOR accounted for the RFLP losses (Fig. 6B). Dramatic alterations occurred within the S+F where five of six parental NORs were retained and new rDNA RFLPs were created and amplified differentially among somaclonal-variant plants (Fig. 6C). The molecular basis of the new RFLPs involved increased numbers of a 340-bp subrepeating element within the rDNA IGS suggesting that recurrent cycles of unequal recombination occurred at high frequency within the rDNA in somatic lineages.Differences between the parental lines were also detected for the amplification level of another tandemly-repeated sequence (C300) that was cloned in M.coerulea ( Calderini et al., 1997) and found to be interspersed in the genome of the M.sativa-M.coerulea-M.falcata complex. In general C300 was most abundant in M.coerulea with 3x105 copies per haploid genome, a value almost twice that found in M.sativa, even though the latter has a double chromosome complement. Conversely, the number of copies was markedly lower in M.falcata (0.8X105) and this sequence was totally absent in M.arborea. There was also variation in the C300 copy number in somatic hybrids which was dependent on the contribution of each parent. The copy number of S+F was very similar to that of M.sativa, indicating that most chromosomes (probably 32 of 33) were derived from that parent. The lower number of copies in S+A than in M.sativa is consistent with the observation that this hybrid lacked some M.sativa chromosomes (Nenz et al., 1996). Although both parental chromosome complements were present in S+C, the copy number recorded in this hybrid was lower than the expected one. The species-specificity of C300 in M.sativa compared to M.arborea was used in FISH (
Fig. 7A-B) experiments to reveal the genome contribution of M.sativa in S+A. In this plant, the C300 probe hybridised to 26 out of 56 chromosomes, indicating incomplete integration of both parental genomes in the hybrid. In fact, if M.sativa is considered to have a maximum of three chromosomes without signal, a maximum of 29 chromosomes would be accounted to M.sativa and a minimum of 27 to M.arborea. Therefore, no dramatic imbalance of the parental chromosome complements in this somatic hybrid was observed.Both Southern and cytological analyses of tandemly-repeated sequences (rDNA and C300) substantially confirmed the analyses carried out with RAPD and RFLP indicating stability or small-scale rearrangements in S+C and large-scale genomic rearrangements in both S+A and S+F.
c) Morphological analyses
To investigate how the genome rearrangements detected at cytological and molecular levels could have affected the field performance of somatic hybrids, a comparative morphological analysis of traits related to forage production was carried out. In general S+C and S+F showed a morphological resemblance to M.sativa while S+A appeared intermediate between the parents. The most important requirement for an alfalfa variety is high forage yield. Our hexaploid somatic hybrids S+C, despite a high degree of heterozygosity, a high ploidy level and high values of stem diameter, width, length and weight of leaves, did not outperform the tetraploid parent in leaf and stem weight, the traits most correlated to forage production. The somatic hybrids showed a forage production per plant significantly higher than the diploid parent and similar to that of M.sativa (Pupilli et al., 1995). The two species M.sativa and M.arborea differ very distinctly in many morphological traits and the resulting hybrids display a characteristic intermediate plant morphology with a rather vigorous aspect, a lower number of stems per plant and a slightly higher leafiness compared to M.sativa. The number of stomata was intermediate between the parents and this could be a very important feature since the high stomata number of M.sativa compared to M.arborea could be related to the adaptation potential of this shrub species to dry environments (Nenz et al., 1996). However, the most striking feature of S+A was the inability of these plants to flower. As indicated by molecular and cytological data M.sativa contribution was prevalent in S+F, and hybrids resulted highly similar to M.sativa for most of the traits analysed.
The application of somatic hybridisation to plant breeding has been limited in all the experiments performed by the low number of hybrid plants derived from different fusion events: in fact chromosome loss that often occurs in somatic hybrids is not related to the duration of in vitro culture but to the status of protoplasts before and after electrofusion. Therefore an effort for optimising large scale production of somatic hybrids needs still to be made to select within a large hybrid population the individuals carrying the most useful gene combinations. In fact, through this technique it is possible to transfer simultaneously multigenic traits but, for this purpose it is necessary to select for the right allele combination within a large population of recombinants. Besides, if somatic hybridisation has to be used to enrich the cultivated germplasm of useful genes, the greatest part of the work is the screening for genotypes having these requisites. Somatic hybridisation is a very difficult job since the result of an experiment is a well defined product represented by a living plant. However, the method is a useful tool for remodelling the entire architecture but is poor or ineffective for transferring specific oligogenic traits. In this respect, the experiment carried out by Li et al. (1993) is a case in point. They attempted to transfer from sainfoin the genes coding for the synthesis of condensed tannins. For this purpose they produced a great many asymmetric somatic hybrids but none possessing the requested features. On this point genetic transformation has evident superiority.
Genetic transformation in alfalfa has been attempted since the mid-eighties using reporter genes and Agrobacterium tumefaciens and A.rizhogenes in order to set up the optimal method of producing transgenic plants in alfalfa. Notwithstanding the long time passed and the great amount of work done, no great progress has been achieved. In fact, although transgenic alfalfa plants have been developed by different methods and a large number of genes have been introduced, the number of transgenic plants produced per experiment remains so extremely low that it negatively affects the possibility of selecting the best transgenic plants, where copy number, genome position and interactions with the endogenous sequences are optimised for the purposes of the experiment.
The most serious disturbance in the production of transgenic alfalfa is the regeneration of some untransformed plants that have escaped screening during in vitro selection. Different selective agents were tested in our lab in order to identify the most suitable one, but none better than kanamycin was found, although kanamycin is not a reliable selective agent at the callus stage in alfalfa.
The problem of the production of false transformants was addressed using low amounts of antibiotic during embryogenesis, the most critical phase of tissue culture. However, to regenerate large quantities of plants it is necessary to grow a very large number of calli and no antibiotic treatment could prevent a number of regenerants escaping selection. In a recent experiment conducted in our lab, it was estimated that 33% of false transformants were obtained with both callus growth and embryogenesis carried out on medium containing 20 mg/l kanamycin. This means that, after regeneration, PCR analysis of the putative transgenic plants is indispensable.
These problems could be partially overcome by using Agrobacterium rhizogenes. With this vector transgenic tissues are easily identified, thereby reducing the problem of the regeneration of false transformants. When binary A.rhizogenes vectors are employed it may happen that a hairy root be transformed only with Ri-T-DNA. In this case the transgenic tissue is useless but this only happens on average in 30% of cases (Damiani and Arcioni, 1991; Damiani et al., 1993), therefore escapes are limited in number and can be screened out early if a colour-producing gene, such as uidA, is used as reporter. The introduction of wild type Ri DNA is however of controversial use, as these genes deeply affect plant structure and physiology and may have positive as well as negative effects. Hairy root phenotypes are surely more appealing for forage production, as they feature increased leafiness and reduced stem dimensions (
Fig. 8A). In contrast, the modification of the root apparatus, which turns fibrous and shallow instead of deep and tap-rooted (Fig. 8B), is of dubious utility (Spanò et al., 1987).Rol genes, which constitute most of the Ri DNA, also have a positive effect on pathogen resistance (Flandung and Gieffers, 1993), but the most strikingly negative effect of hairy root transformation is reduced persistence, in fact many transgenic plants were observed to persist no more than one year (Damiani and Paolocci, 1991).
The technical problems involved in transformation should not however restrain the application of this technique of gene introduction for useful traits, because once the gene of interest has been isolated and the construct with the most appropriate promoter has been made, the extra work necessary for producing the optimal number of transgenic plants is not a limiting factor.
The possibility of successfully introducing genes from alien species is currently being investigated in our lab, with special emphasis on the problems related to alfalfa forage quality. A dual approach is followed: improvement of protein composition and the introduction of moderate amounts of condensed tannins that may prevent bloating and increase protein assimilation.
Various research projects are concerned with the balance of protein content in legume species through genetic engineering. A chimaeric gene encoding the methionine-rich 2S albumin of the Brazil nut has been stably expressed and inherited in transgenic grain legumes (Saalbach et al., 1994). As regards the nutritional improvement of forage legumes,
d -zein, a sulphur amino acid-rich protein, has been expressed in transgenic white clover (Sharma et al., 1998), as well as a sulphur-rich seed albumin from sunflower accumulated in the leaves of both transgenic subterranean clover (Khan et al., 1996) and transgenic alfalfa (Tabe et al., 1995). The use of genes coding for sulphur-rich proteins in the transformation of forage legumes followed evidence that sulphur-containing amino acids have an important role in the efficiency of dairy, meat and wool production (Barry, 1981; Reis, 1979; Rogers et al., 1979). In the case of sheep, it is also well established that unprotected dietary proteins are rapidly degraded by bacteria in the rumen and converted into microbial proteins. Thus, the rumen microbial proteins are the main source of protein for animals, but they are low in sulphur amino acids and, as a result, there is a significant loss of cysteine and methionine during the conversion of ingested protein into microbial protein (Thompson, 1982; Khan et al., 1996). For this reason, the sulphur-rich protein candidate for expression in transgenic forage legumes should be resistant to proteolytic degradation in the rumen. With the long-term goal of improving alfalfa forage quality, stable expression of a zein transgene has been attempted in alfalfa, as well as in the model species birdsfoot trefoil and tobacco (Bellucci et al., 1997; Bellucci et al., 1998; Bellucci et al., 1999). It makes sense to verify the expression of a gene construct in a model species rather than transform the species of interest with the gene construct. In fact, it may happen that problems related to poor accumulation of heterologous proteins in transgenic plants make it necessary to modify the original gene construct in order to have a high protein expression. Wandelt et al., (1991) achieved only low levels of ovalbumin protein translation in transgenic alfalfa and tobacco leaves. Bagga et al., (1992) transformed alfalfa with a b -phaseolin gene under control of the 35S promoter. Although all tissues examined contained high levels of phaseolin transcripts, only mature seeds showed significant accumulation of the protein. Furthermore, Wandelt et al., (1992) reported that vicilin, the pea seed storage protein, was rapidly degraded in the leaf vacuoles of transgenic alfalfa.We decided to transform alfalfa with genes coding for zein proteins because zeins are the major protein reserves in maize kernels and detailed information is available on the structure and physiology of this kind of prolamins (Shewry et al., 1995). Zeins, encoded by a multigene family (Hagen and Rubenstein, 1980), can be classified into different relative mobility (Mr) groups,
a -, b -, d - and g -zeins (Wilson, 1991). b -, d - and g -zeins are rich in the essential sulphur amino acids methionine and cysteine. Moreover, zeins are usually rumen-protected (Bagga et al., 1995; Sharma et al., 1998). Thus, genes coding for zeins are extensively utilised in genetic engineering programmes (Bellucci et al., 1997; Bellucci et al., 1998; Bellucci et al., 1999; Bagga et al., 1995; Bagga et al., 1997; Sharma et al., 1998). These studies indicated that transformed plants accumulate zeins in vegetative tissues and open up the possibility of using zeins to increase the sulphur-rich amino acid content in edible tissues of forage plants.We are currently engaged in transforming forage legumes and tobacco with two cDNAs, one coding for a
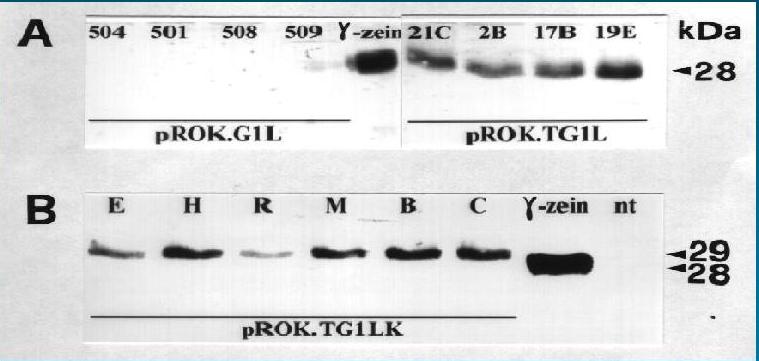
g -zein and another one for a b -zein. Initially, we analysed the expression of the g -zein cDNA (G1L), driven by CaMV 35S or by ribulose bisphosphate carboxylase small subunit (rbcS) promoter, in tobacco and in the two forage species, alfalfa and birdsfoot trefoil (Bellucci et al., 1997). Although G1L transcripts were detected in all transgenic plants with both promoters, g -zein only accumulated in tobacco tissues in very low quantities (0.05 % of alcohol-soluble proteins, Fig. 9A). The lack of any significant accumulation of g -zein in any of the species investigated was ascribed to post-transcriptional effects: 5’UTR of G1L cDNA was indicated as probably responsible for either mRNA instability or difficulty of in vivo translation. To ascertain whether this DNA sequence had been an impediment to g -zein accumulation, we generated another gene construct (pROK.TG1L) in which the 5’UTR of G1L was replaced with the untranslated leader from the coat protein mRNA of tobacco mosaic virus (TMV). In this new chimaeric gene G1L was placed under control of the rbcS promoter in order to avoid using the 35S promoter, frequently associated with gene silencing effects. In a previous study, no G1L mRNA had been detected in 35S-G1L birdsfoot trefoil plants, due to a transcriptional block (Bellucci et al., 1999). pROK.TG1L transformants accumulated high levels of g -zein (Fig. 9A), in alfalfa, in birdsfoot trefoil and in tobacco (expression of g -zein in tobacco transformants ranged between 0.022 - 0.62 % of total water-soluble protein). This suggested that TMV leader enhanced translation of G1L mRNA and that the mRNA translation efficiency was a rate-limiting step for G1L protein expression.To further increase
g -zein accumulation in the transgenics, a g -zein mutant was constructed by adding a KDEL motif, a retention signal described for ER resident proteins, before the stop codon. This gene construct (pROK.TG1LK) was identical to the plasmid pROK.TG1L, except for the presence of the amino acid sequence TSEKDEL. pROK.TG1LK was also used to transform alfalfa, birdsfoot trefoil and tobacco. In maize, zeins accumulate in protein bodies arising from the rough endoplasmic reticulum (ER) (Larkins and Hurkman,1978) but, unlike other ER-resident proteins they do not possess the canonical (K/H)DEL ER retention sequence, so there must be other factors preventing zeins from exiting the ER and allowing their deposition in protein bodies. Geli et al., (1994) transformed Arabidopsis thaliana with a g -zein coding sequence and the protein accumulated in leaves and roots. g -zein was retained in ER-derived protein bodies and a proline-rich repeat domain was indicated as necessary for g -zein retention in the ER.Our hypothesis was that the addition of the ER retention signal KDEL might further enhance
g -zein accumulation by increasing the percentage of protein molecules retained in the ER. g -zein:KDEL also was highly expressed in alfalfa, birdsfoot trefoil and tobacco (expression of g -zein in tobacco transformants ranged between 0.011-0.76% (Fig. 9B) of total water-soluble protein). Apparently our hypothesis was not correct because plants transformed with either pROK.TG1L or pROK.TG1LK accumulated similar quantities of g -zein (Bellucci et al., 1998). However, statistical analyses carried out on 28 tobacco transformants by quantitative western blots demonstrated that the addition of the KDEL retention signal favoured production of g -zein:KDEL rather than the native g -zein of pROK.TG1L transformants (average values were 0.24 and 0.12% of the water-soluble protein, respectively).Recent studies have shown that nascent prolamins associate transiently in an ATP-dependent manner with BiP upon insertion into the ER (Li et al., 1993; Galili et al., 1995). Moreover, evidence for a possible role of BiP in prolamin assembly has come to light from observing increased BiP accumulation in maize mutants and transgenic tobacco plants overproducing
b - and d -zeins (Boston et al., 1991; Bagga et al., 1995). In pROK.TG1L and pROK.TG1LK transformants BiP was also increased. We measured BiP accumulation in the tobacco plants transformed with these two gene constructs and in ten wild-type plants as controls. Although pROK.TG1LK transformants had the higher level of g -zein accumulation, they increased BiP accumulation less than pROK.TG1L transformants. These results suggest that the two-fold increase in the level of g -zein:KDEL could probably be attributed to an improvement in the ER retention mechanism of zeins in the transgenic tobacco cells due to the addition of the KDEL ER retention signal. Furthermore, it is reasonable to assume that the g -zein:KDEL protein is less dependent on the chaperone-like assistance of BiP because the g -zein mutant has the same ER retention signal as BiP.Up to now, preliminary results have been obtained with the
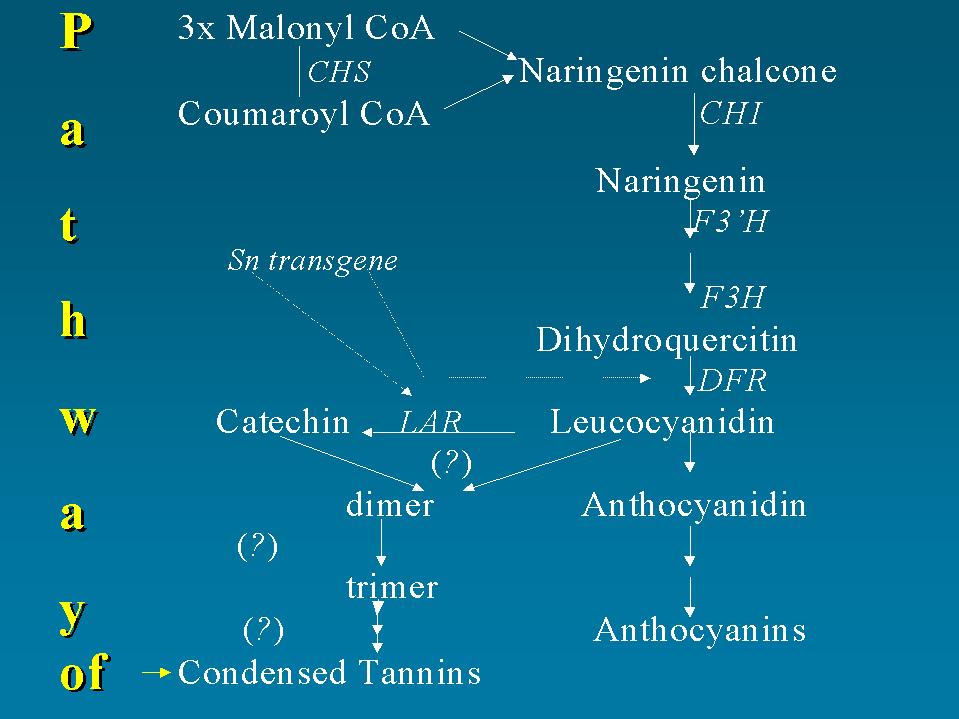
b -zein:KDEL. One gene construct was assembled in which the mutated b -zein cDNA (G2) was located between the left and right borders together with other two genes, nptII and uidA. This allowed us to select the transformed plants using both kanamycin in the tissue culture medium and GUS assay to individuate plants expressing the foreign genes. Transformed tobacco plants accumulated b -zein:KDEL up to 1% of water-soluble protein and we are now regenerating alfalfa plantlets from transformed calli. Our objective was to coexpress g -zein with b -zein in alfalfa to improve the expression of sulphur-rich zeins. In tobacco, we already obtained plants that accumulated both types of zeins in the leaves by crossing plants expressing g - or b -zein.A study of the prevention of protein degradation was undertaken to complement the work described above. It is useless to improve a forage by introducing superior proteins if most of these are degraded and wasted when introduced into the rumen. In this connection it has been demonstrated that moderate amounts of condensed tannins (CT) in the forage prevent protein degradation (Tanner et al., 1995) and consequently increase the nutritive value of the forage. CT bind to protein and make them unavailable to the degrading action of rumen flora. Thanks to the same mechanism, forage containing CT does not cause bloating, a deadly condition affecting ruminants fed on fresh forage legumes. On the other hand, an excess of CT in the forage has negative consequences, as their inhibiting action on salivary enzymes gives a bitter taste to the forage and animals refuse to graze. Thus, it would be useful to be able to modulate the CT level in forage legumes. In some species excess CT should be reduced while in others, such as those cultivated the most extensively (alfalfa and clovers), CT are absent from the edible tissue and therefore their synthesis should be induced. The possibility of altering this trait is hampered by insufficient knowledge of the genetic control of their synthesis. CT are secondary metabolites originating from a branch of the anthocyanin pathway. For this reason their biosynthetic pathway is well characterised and the genes coding for the different enzymes have been cloned in different species (Antirrhinum, Sommer and Saedler, 1986; Petunia, Koes et al., 1989; maize, Wienand et al., 1986; Vitis, Sparvoli et al., 1994), up to the last common intermediate. The pathway is trans-activated by some regulator genes belonging to the myc and myb families (van der Meer et al., 1993) which interact with promoter regions of the structural genes and initiate their transcriptions. The biosynthesis of anthocyanidin and CT share the same sequence of reactions to the last common intermediate, leucoanthocyanidin, the formation of which is catalysed by the enzyme dihydroflavanol reductase (DFR,
Fig. 10). Little is known about the molecular biology of the branch of the flavonoid pathway leading from leucoanthocyanidin to CT. The NADPH-dependent reduction of labelled leucoanthocyanidin to catechin has been demonstrated in a number of species, including L. corniculatus (Tanner and Kristiansen, 1993; Tanner, unpublished). This activity is due to a leucoanthocyanidin reductase (LAR, Fig. 10). The nature of the subsequent condensation steps leading to CT is unknown.In order to utilise genetic transformation for manipulating CT synthesis a prerequisite is the availability of the gene sequences coding for the LAR enzyme and for the monomer-condensing enzymes, if existing. For this purpose different approaches are being taken in various laboratories. One is a protein-based method consisting in isolating the LAR enzyme and utilising its amino acid sequence for determining the nucleotidic one. Other approaches are based on the production of mutants through modification of the DNA sequence, like the transposon tagging method, or through modification of gene expression with an antisense strategy or cosuppression.
We have adopted this latter strategy and obtained very promising results (Damiani et al., 1999). Working on Lotus corniculatus as gene donor species, we transformed various genotypes, characterised by contrasting levels of CT (very low, low, high) with the maize gene Sn. This gene is a trans-activator of the R family and determines a strong red pigmentation in different tissues. We only used its coding region under a constitutive promoter (35S) and easily produced three populations of transformants, and because each population was derived from a well characterised plant, all the plants of the same population were isogenic, the only differences among them being the number of copies of the transgene and its position in the genome. These minor differences were, however, very significant in the plant phenotype: some plants were totally suppressed for tannin synthesis in leaves, others, on the contrary, were elicited for this trait (
Fig. 11). CT content of roots also was modified (Fig. 12) and recent observations indicated that plants with null CT levels in leaves and roots are also poor in rooting while plants with high levels of CT rooted profusely. A possible explanation for this may be found in the transgene-endogene interaction theory. It has been demonstrated that in some cases where a transgene has homology with an endogenous gene, the two of them can negatively interact and no mRNA of either is produced. The mechanism of the silencing has not been demonstrated yet, but there is plenty of evidence suggesting that an excess of homologous RNAs may be the cause (Metzlaff et al., 1997). We adopted this view to account for the presence of null CT phenotypes, while we explained the elicited phenotypes by a mere additive action of mRNAs from the transgene and the endogene. These assumptions were supported by several observations (Paolocci et al., 1999):1) suppressed plants had several copies of the transgene while unsuppressed plants had only one;
2) transgene expression was detected in elicited plants but not in suppressed plants (
Fig. 13);3) the expression of a Lotus myc-like gene, isolated through RT-PCR with degenerated primers deduced by the Sn sequence, followed the same pattern of expression as the transgene (
Fig. 13);4) the expression of dfr and the relative enzymatic activity were enhanced in CT elicited transformants, but very poor or null in suppressed plants (
Fig. 13).The newly produced genotypes will be exploited for isolating all the suppressed genes of the pathway, including those still unknown which are responsible for LAR activity and for the subsequent condensation of catechin.
References
Arcioni, S., E. Nenz, F. Pupilli, and F. Damiani. 1994. Gene transfer into Medicago through somatic hybridization. Report of the thirty-fourth North American Alfalfa Improvement Conference, July 10-14, 1994 Guelph, Canada pp. 45.
Arcioni, S., F. Pupilli, M. Pezzotti, E. Falistocco, and F. Damiani. 1987. Interspecific hybrid lines of M.sativa and M.arborea by protoplast electrofusion p.259-260. In K.J. Puite, J.J.M. Dons, H.J. Huizing, A.J. Kool, M. Koornneef, and F.A. Krenes (ed.) Progress in Plant Protoplasts Research. Kluwer Academic Publishers, Dordrecht.
Bagga, S., D. Sutton, J.D. Kemp, and C. Sengupta-Gopalan. 1992. Constitutive expression of the
Bagga, S., H. Adams, J.D. Kemp, and C. Sengupta-Gopalan. 1995. Accumulation of a 15-Kilodalton zein in novel protein bodies in transgenic tobacco. Plant Physiol. 107: 13-23.
Bagga, S., H. Adams, F.D. Rodriguez, J.D. Kemp, and C. Sengupta-Gopalan. 1997. Coexpression of the maize
d -zein and b -zein genes results in stable accumulation of d -zein in endoplasmic reticulum-derived protein bodies formed by b -zein. Plant Cell 9: 1683-1696.Barry, T., 1981. Protein metabolism in growing lambs fed on fresh ryegrass (Lolium perenne) white clover (Trifolium repens) pasture ad lib.. British Journal Nutrition 46:521-532.
Bellucci, M., A. Alpini, F. Paolocci, F. Damiani, and S. Arcioni. 1999. Transcription of a maize cDNA in Lotus corniculatus is regulated by T-DNA methylation and transgene copy number. Theor. Appl. Genet. 98: 257-264
Bellucci, M., B. Lazzari, A. Viotti, and S. Arcioni. 1997. Differential expression of a
g -zein gene in Medicago sativa, Lotus corniculatus and Nicotiana tabacum. Plant Sci. 127:161-169.Bellucci, M., F. Paolocci, A. Alpini, and Arcioni S. 1998. A biotechnological approach to increase the content of sulphur-rich proteins in forage leguminous species. Proceedings of the 17th General Meeting of the European Grassland Federation, G. Nagy and K. Peto (ed.), Debrecen Agricultural University, Debrecen, Hungary, May 18-21, 219-221.
Boston, R.S., E.B.P. Fontes, B.B. Shank, R.L. Wrobel. 1991. Increased expression of the maize immunoglobulin binding protein homolog b70 in three zein regulatory mutants. Plant Cell 3: 497-505.
Brummer, E.C., J.H. Bouton, and G. Kochert. 1993. Development of an RFLP map in diploid alfalfa. Theor. Appl. Genet. 86: 329-332.
Calderini, O., F. Pupilli, F. Paolocci, and S. Arcioni. 1997. A repetitive and species-specific sequence as a tool for detecting the genome contribution in somatic hybrids of the genus Medicago. Theor. Appl. Genet. 95:734-740.
Damiani, F., F. Paolocci, P.D. Cluster, S. Arcioni, G.J. Tanner, R.G. Joseph, Y.G. Li, J. DeMajnik, and P.J. Larkin. 1999. The maize Sn regulatory transgene alters proanthocyanidin production in Lotus corniculatus in some tissues by transactivation and in others by co-suppression of an endogenous regulator. Aust. J. Plant Physiology 26: 159-169.
Damiani, F., and F. Paolocci. 1991. Effetto della trasformazione con plasmide Ri su caratteri morfologici e fisiologici in leguminose foraggere. Atti XXXV Convegno Annuale della Società Italiana di Genetica Agraria. Pisa 23-26 Settembre pp. 85-86.
Damiani, F., and S. Arcioni. 1991. Transformation of Medicago sativa L. with an Agrobacterium rhizogenes binary vector carrying the hygromycin resistance gene. Plant Cell Reports 10:300-303.
Damiani, F., E. Nenz, F. Paolocci, and S. Arcioni. 1993. Introduction of hygromycin resistance in Lotus spp, through Agrobacterium rhizogenes transformation. Transgenic Res. 2:330-335.
Damiani, F., M. Pezotti, and S. Arcioni. 1988. Electric field mediated fusion of protoplasts of Medicago sativa L. and Medicago arborea L.. Journal of Plant Physiology 132: 474-479.
Flandung, M., and W. Gieffers. 1993. Resistance reactions of leaves and tubers of rolC transgenic tetraploid potato to bacterial and fungal pathogens. Correlation with sugar, starch and clorophyll content. Physiological and Molecular Plant Pathology 42:123-132.
Galili, G., S. Giorini-Silfen, Y. Shimoni, Y. Altschuler, H. Levanony, N. Shani, H. Karchi, and E. Galun. 1995. Assembly and intracellular transport of wheat storage proteins. J. Biol. Chem. (Suppl 19A-019): 132.
Geli, M.I., M. Torrent, D. Ludevid. 1994. Two structural domains mediate two sequential events in zein targeting: protein endoplasmic reticulum retention and protein body formation, Plant Cell 6: 1911-1922.
Hagen, G., and I. Rubenstein. 1980. Complex organization of zein genes in maize. Gene 13: 239-249.
Khan, M.R.I., A. Ceriotti, L. Tabe, A. Aryan, W. McNabb, A. Moore, S. Craig, D. Spencer, and T.J.V. Higgins. 1996. Accumulation of a sulphur-rich seed albumin from sunflower in the leaves of transgenic subterraneum clover (Trifolium subterraneum L.). Transgenic Res. 5: 179-185.
Koes, R.E., C.E. Spelt, P.J.M. van den Elzen, and J.N.M. Mol. 1989. Cloning and molecular characterization of the chalcone synthase multigene family of petunia. Gene 81:245-257.
Larkins, B.A., and W.J. Hurkman. 1978. Synthesis and deposition of zein in protein bodies of maize endosperm. Plant Physiol. 62: 256-263.
Li, X., Y. Wu, D.Z. Zhang, J.W. Gillilin, R.S. Boston, V.R. Franceschi, and T.W.B. Okita. 1993. Rice prolamin protein body biogenesis: A BiP-mediated process. Science 262: 1054-1056.
Li, Y.G., G.J. Tanner, A.C. Delves, and P.J. Larkin. 1993. Asymmetric somatic hybrid plants between Medicago sativa L. (alfalfa, lucerne) and Onobrychis viciifolia Scop. (sainfoin). Theor. Appl. Genet. 87, 455-463.
Long, E.O. and David, I.B. 1980. Repeated genes in eukaryotes. Annual Review Biochemistry 49:727-764.
MacCoy, T.J. and E.T. Bingham. 1991. Alfalfa cytogenetics p.399-418. In T. Tsuchiya and P.K. Gupta. (ed.). Chromosome engineering in plants: genetics, breeding, evolution part B. Elsevier Publ. Amsterdam Oxford, New York, Tokyo.
McCoy, T.J. and E.T. Bingham. 1988. Cytology and cytogenetics of alfalfa p737-776. In Hanson, A.A., D.K. Barnes and R.R. Hill, (ed.) Alfalfa and Alfalfa Improvement . Agronomy Monograph N. 29 ASA-CSSA-S, Madison, Wisconsin, USA.
Metzlaff, M., M. O'Dell, P.D. Cluster, and R.B. Flavell. 1997. RNA-mediated RNA degradation and chalcone synthase A silencing in petunia. Cell 88:845-854.
Nenz, E., F. Pupilli, F. Damiani, and S. Arcioni. 1996. Somatic hybrid plants between the forage legumes Medicago sativa L. and Medicago arborea L.. Theor. Appl. Genet. 93:183-189.
Paolocci, F., R. Capucci, S. Arcioni, and F. Damiani. 1999. Birdsfoot trefoil: a model for studying the synthesis of condensed tannins. In Groos G.G., R.W. Heminghway, and T. Yoshida, (ed.): Plant Polyphenols 2. Chemistry and Biology. Plenum press (in press).
Pupilli, F., S. Arcioni, F. Damiani, and M. Pezzotti. 1991. Protoplasts in the genus Medicago and isoenzyme analysis of parental and somatic hybrid cell lines. Plant Breeding 106:122-131.
Pupilli, F., Businelli, S., Caceres, M.E., Damiani, F. and Arcioni, S. 1995. Molecular, cytological and morpho-agronomical characterization of hexaploid somatic hybrids in Medicago. Theor. Appl. Genet. 90:347-355.
Pupilli, F., G.M. Scarpa, F. Damiani, and S. Arcioni. 1992. Interspecific somatic hybrid plants in the genus Medicago developed through protoplast fusion. Theor. Appl. Genet. 84:792-797.
Reis, P.J. 1979. Effects of amino acids on the growth and properties of wool p. 233-242. In J.L. Black and P.J. Reis (ed.) Physiological and environmental limitations to wool growth, University of New England Publishing Unit, Armidale, NSW, Australia.
Rogers, G., A. Bryant, A., and L. McLeay. 1979. Silage and dairy cow production. III Abomasal infusions of casein, methionine and glucose and milk yield and composition. N.Z.J. Agricultural Research 22:533-541.
Rogers, S.O. and A.J. Benedich. 1987. Ribosomal RNA genes in plants: variability in copy number and in the intergenic spacer. Plant Mol. Biol. 9:509-520.
Saalbach, I., T. Pickardt, F. Machemehl, O. Saalbach, O. Schieder, and K. Muntz. 1994. A chimeric gene encoding the methionine-rich 2S albumin of the Brazil nut (Bertholletia excelsa H.B.K.) is stably expressed and inherited in transgenic grain legumes. Mol. Gen. Genet. 242: 226-236.
Sharma, S.B., K.R. Hancock, P.M. Ealing, D.W.R. White. 1998. Expression of a sulphur-rich maize seed storage protein,
d -zein, in white clover (Trifolium repens) to improve forage quality. Molecular Breeding 4:435-448.Shewry, P.R., J.A. Napier, A.S. Tatham. 1995. Seed storage proteins: structures and biosynthesis. Plant Cell 7: 945-956.
Sommer, H., and H. Saedler. 1986. Structure of the chalcone synthase gene in of Antirrhinum majus. Mol. Gen. Genet. 202:429-434.
Spanò, D., D. Mariotti, M. Pezzotti, F. Damiani, and S. Arcioni. 1987. Hairy root transformation in alfalfa (Medicago sativa L.). Theor. Appl. Genet. 73:523-530.
Sparvoli, F., C. Martin, A. Scienza, G. Gavazzi, and C. Tonelli. 1994. Cloning and molecular analysis of structural genes involved in flavonoid and stilbene biosynthesis in grape (Vitis vinifera L.) Plant Mol. Biol. 24:743-755.
Tabe, L., T. Wardley-Richardson, A. Ceriotti, A. Aryan, W. McNabb, A. Moore, and T.J.V. Higgins. 1995. A biotechnological approach to improving the nutritive value of alfalfa. Journal Animal Science 73:2752-2759.
Tanner, G.J. and K.N. Kristiansen. 1993. Synthesis of 3,4-cis-leucocyanidin and enzymatic reduction to catechin. Anal. Biochem. 209:274-277.
Tanner, G.J., P. Moate, L. Dailey, R. Laby, P.J. Larkin. 1995. Proanthocyanidins (condensed tannins) destabilise plant protein foams in a dose dependent manner. Aust. J. Agric. Res. 46:1101-1109.
Thompson, D.J. 1982. The nitrogen supplied by and the supplementation of fresh or grazed forage p. 53-66. In Thompson, D.J., E. Beever, and R.G. Gunn (ed.) Forage Protein in Ruminant Animal Production, Occasional Publication N. 6, British Society of Animal Production.
van der Meer, I.M., A.R. Stuitje, and J.N.M. Mol. 1993. Regulation of general phenyl-propanoid and flavonoid gene expression p 125-155. In Verma, D.P.S. (ed.) Control of plant gene expression, CRC Press, Boca Raton, FL.
Wandelt, C.I., M.R.I. Khan, S. Craig, H.E. Schroeder, D. Spencer, and T.J.V. Higgins. 1992. Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulated to high levels in the leaves of transgenic plants. Plant J. 2: 181-192.
Wandelt, C.I., W. Knibb, H.E. Schroeder, M.R.I. Khan, D. Spencer, S. Craig, T.J.V. Higgins. 1991. The expression of an ovalbumin and a seed protein gene in the leaves of transgenic plants p. 471-478. In Hermann, R.G., and B.A. Larkins. (ed.) Plant molecular biology, 1990 Plenum Press, New York, 1991,.
Wienand, U., U. Weydemann, U. Niesbach-Kloesgen, OP.A. Peterson, and H. Saedler. 1986. Molecular cloning of C2 locus of Zea mays, the gene coding for chalcone synthase. Mol. Gen. Genet. 203:202-207.
Wilson, C.M. 1991. Multiple zeins from maize endosperm characterized by reverse-phase high performance liquid chromatography. Plant Physiol. 95: 777-786.
Figure legends
Fig. 1. Electrofusion between mesophyll (M. sativa) and callus (M.coerulea) protoplasts. A. the two protoplasts are aligned by the AC electric field. B. application of fusion pulse. C. heterokaryon 3 min after fusion pulse. D. heterokaryon 5 min after fusion pulse. E. mesophyll protoplasts observed under fluorescent light. F. callus protoplasts (yellow-green colour) treated during enzymatic digestion with fluorescein isothiocyanate mixed with mesophyll protoplasts (red colour) under fluorescent light. G. heterokaryon under fluorescent light.
Fig. 2. Cytological analysis of somatic hybrids M.sativa + M.coerulea. A. metaphase I with laggards (black dashes). B. anaphase II with laggards (black dashes).
Fig. 3. Metaphase chromosomes of a somatic hybrid plant M.sativa + M.falcata. Arrows indicate two chromosomes of unexpected length, probably a consequence of a translocation phenomenon.
Fig. 4. Hybridising banding patterns of the probe 1H11 with the DNA digests of M.sativa (s), M.coerulea (c) and 15 somatic hybrids (h1..15). The restriction enzyme was AluI. Arrow indicates the M.coerulea-specific band missing in the hybrid patterns and the molecular weights (m) are indicated in base pairs.
Fig. 5. A. RAPD analysis of somatic hybrid plants (sf1-sf3) M.sativa + M.falcata with the random primer RP7. B. Southern analysis of the RAPD from RP7 (probe 530 bp M.falcata-specific RP7 fragment). Somaclones of M.falcata (s1-s3) and M.sativa (f1-f3) parental plants were analysed.
Fig. 6. RFLP pattern of rDNA intergenic spacers (IGS). A. IGS RFLP patterns from M.sativa (s), M.coerulea (c) and their hybrids (SC1-SC3) with the ribosomal gene 18S as probe. B. IGS RFLP pattern from M.sativa (s), M. arborea (a) and their hybrids (SA2-SA4) with the ribosomal 25S as probe. C. IGS RFLP from M. sativa (s), M. falcata (f) and their hybrids (SF3-SF10) with the probe 25S.
Fig. 7. A-B. Fluorescent in situ hybridisation to somatic metaphase of M.sativa and the somatic hybrid M.sativa + M. arborea. The chromosomes were stained with the fluorochrome DAPI (blue) and the pink spots correspond to the hybridisation sites of rhodammine-labelled C300. A. M.sativa metaphase in which the solid arrow indicates a single signal close to the centromere, the open arrow indicates two signals in a pericentromeric position, and the arrowhead indicates a pericentromeric and a subtelomeric signal. B. M.sativa + M.arborea metaphase.
Fig. 8. Parts of the same genotype of M. sativa not transformed (left) and transformed with A. rhizogenes (right). A. Stems. B. Roots.
Fig. 9. Western analysis of tobacco transformants with the anti-g -zein antibody. A. four pROK.G1L and four pROK.TG1L transformants. 40 µg of leaf proteins soluble in saline buffer with 4% 2-mercaptoethanol were separated by SDS-PAGE and subjected to western analysis. 0.2 µg of purified g -zein extracted from maize endosperm was used as a standard. B. six pROK.TG1LK transformants and one non-transformed plant. The same analysis as in Figure 9A was performed, except that 0.4 µg of purified g -zein was loaded. nt, non-transformed.
Figure 10. Biosynthetic pathway of condensed tannins. The known enzymes are shown italicised, the putative condensing enzyme are indicated with (?). The interaction between the Sn transgene and the enzymatic activities are indicated with ( ).
Fig. 11. Leaves of elicited (left) and suppressed (right) plants stained for condensed tannins.
Fig. 12. Roots of suppressed (upper portion) and elicited (lower portion) plants stained for condensed tannins.
Fig. 13. Northern analyses of transgenic suppressed (S50-9) and elicited (S50-10) plants derived from the wild genotype S50 (control plant). Probes are indicated. The a elongation factor (EF) was utilized as control of gene expression.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}